导图社区 酶
- 32
- 0
- 0
- 举报
酶
酶学概论,酶是由细胞合成的,在机体内行使催化功能的生物催化剂,有酶催化的反应称为酶促反应,主要是蛋白质,少数是RNA。
编辑于2023-10-23 22:03:25- 酶
- 相似推荐
- 大纲



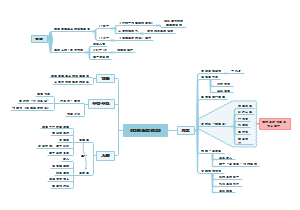

酶
酶的化学本质
酶是由细胞合成的,在机体内行使催化功能的生物催化剂
有酶催化的反应称为酶促反应
底物
在酶催化作用下进行化学反应的物质
产物
酶促反应生成的物质
化学本质
主要是蛋白质,少数是RNA
酶的分类
单纯酶
属于单纯蛋白质的酶
缀合酶
除了蛋白质以外,还结合某些非氨基酸成分(辅助因子)
辅助因子
辅酶
与脱辅酶结合松散,使用透析的方法就可以去除的有机小分子
辅基
以共价键和酶蛋白结合,与脱辅酶结合紧密,不能使用透析或超滤的方法去除的有机小分子
金属离子
酶的催化性质
酶与非酶催化剂的共同性质
只能催化热力学允许的反应
反应完成后本身不被消耗或变化,可以重复使用
对正反应和逆反应的催化作用相同
不改变平衡常数,只加快到平衡的速度或缩短达到平衡的时间
酶特有的催化性质
高效性,专一性
绝对专一性
只作用于一种底物,而不作用于任何其他物质
相对专一性
可作用于一类结构相近的底物
基团专一性
键专一性
立体异构专一性
解释酶专一性三种学说
锁与钥匙学说
诱导契合学说
被广泛认可
三点一线学说
区分对映异构体,解释立体专一
在活性中心与底物结合
活性中心
酶分子上直接与底物结合,并与催化作用直接相关的区域
催化部位
催化能力
结合部位
专一性
多为疏水aa残基,少量亲水aa残基(起催化反应的是亲水aa)
活性中心与底物结构互补,且构象并非固定不变的
反应条件温和,对反应条件敏感,容易失活
具有较高的可调节性
影响酶促反应的因素
酶促反应速率
单位时间内,底物或产物浓度的变化量
mol L/S
影响因素
反应温度
离子强度
质子浓度
影响反应pH
金属离子浓度
可以作为酶的辅因子,从而影响反映速率
反应介质pH
影响酶分子的解离状态
影响底物,辅基或辅酶的解离状态
酶激活剂或酶抑制剂
外因
酶浓度
在一定pH和温度条件下,当底物浓度远远大于酶浓度时,反应速率与酶的浓度呈正比关系
底物浓度
底物浓度的三级反应
底物浓度较低时,呈现一级反应 随着底物浓度的增加,反应表现为混合级反应 底物浓度相当高时,呈现零级反应
酶底物中间物学说
内因
米氏反应动力
描述米氏酶反应速率与底物浓度之间的关系
米氏方程的推导
反应速率为初速率
酶底物复合物处于稳态即ES浓度不发生变化
符合质量作用定律
在稳态时,ES的形成速率与ES的解离速率相等
解读米氏方程
Km是酶反应初速率为Vm的一半是的底物浓度
是酶的特征常数,只与酶的性质有关,与酶的浓度无关
在一定条件下,可以反映酶与底物的亲和力,Km越大,意味着酶与底物的亲和力越低
当酶对底物的Km值小于对产物的Km值,则反应有利于正反应,否则有利于逆反应。
Vmax
理论上的最大反应速率,随着酶浓度的变化而变化
Kcat
酶的催化常数或转换数或周转数,具体指单位时间内,一个酶分子将底物转变为产物的分子总数
kcat=k2=Vmax/Et
可以衡量一个酶促反应的快慢
Kcat/Km
用来衡量酶的催化效率,还可以反应一个酶的完美程度
米氏方程的双重性
当底物浓度很低的时候,即S远小于Km,米氏方程转变为
符合一级动力学
当底物浓度很高的时候,即S远大于Km,米氏方程可转变为
符合零级动力学
米氏酶的双倒数作图
酶抑制剂作用
可逆性抑制剂
以次级键和酶可逆性结合
竞争性抑制剂
抑制剂与底物竞争
非竞争性抑制剂
既能与ES结合,也能与E结合
反竞争性抑制剂
只能与ES结合,使得与活性中心结合的底物将无法再转变为产物
不可逆性抑制剂
以强的化学键与酶不可逆结合,一旦失活不可逆转
基因特异性抑制剂
共价修饰酶活性中心上的必需侧链基团
底物类似物抑制剂
结构上相似于底物,在活性中心与酶结合,不可逆的修饰酶活性中心上的必需基团
过渡态类似物抑制剂
与酶促反应的过渡态极为相似,与酶的活性中心进行结合,导致底物无法与酶结合
自杀型抑制剂
受到酶的催化,发生几步反应,但并不形成产物,专而修饰酶的必需基团导致酶活丧失
别构酶的动力学
别构效应
酶分子的非催化部位与某些化合物可逆的,非共价结合后,发生构象的改变,进而改变酶的活性状态,称为酶的别构效应
具有别够效应的酶称为别构酶
能使酶分子发生别构作用的物质称为别构效应物
别构酶的性质
速率,底物浓度曲线为S型
具有别构效应物
对竞争性抑制表现双向反应
低浓度下,诱发别构酶正协同效应,提高底物的反应速率
高浓度下,占据酶的活性中心,底物与S无法正常结合,抑制反应速率
温和变性可以导致别构效应丧失
通常是寡聚酶,且别构酶占少数
Hill方程
h=1,则不是别构酶,没有底物协同性
双曲线
h>1,速率对底物浓度作图呈S型,具有正底物协同性
S型曲线
酶对环境中底物浓度变化敏感,对代谢进行灵敏的调节
h<1,酶具有负底物协同性
表观双曲线
酶对环境中底物浓度变化极度不敏感,保证重要反应不受底物影响,始终进行下去
酶的催化机制
过渡态稳定学说内容
在任何一个化学反应的过程中,反应物需要达到一个特定的高能状态 才能发生反应,这种不稳定的高能状态被称为过渡态,达到过渡态要求反应物有足够的能量以突破活化能
酶与过渡态的亲和力要远远高于对底物(基态)的亲和力,酶的催化作用来源于对过渡的稳定作用
过渡态稳定的化学机制
邻近定向反应
指两种或两种以上的底物同时在酶的活性中心与酶结合,冻结了某些化学键的平动和转动,从而使其采取正确的空间取向,大大提高了底物的浓度
广义的酸碱催化
指酶的活性中心参与催化的氨基酸残基得到或失去质子(转移到过渡态中间物从而达到稳定过渡态的效果。),这种机制参与绝大多数酶的催化过程
质子供体
质子受体
如果pKa的值接近7,该侧链基团可能是最有效的广义酸碱催化剂,如His残基的咪唑基,在生理条件下,一半为酸,一半为碱,释放质子的速率快
反应速率取决于pH以及缓冲液浓度
静电催化
活性中心电荷的分布用来稳定酶促反应的过渡态,酶使用自身带电基团去中和一个反应过渡态形成时产生的相反电荷而进行催化
有时也会通过酶和底物的静电作用将底物引入酶的活性中心
金属催化
金属酶
含有紧密结合的金属离子
金属激活酶
与溶液中的金属离子松散的结合
金属离子的催化方式
作为路易斯酸发挥作用
与带负电的底物结合,促进底物在反应中正确定向
参与静电催化,稳定带负电的过渡态
通过价态的可逆变化,作为电子供体或电子受体参与氧化还原反应
作为酶结构的组成部分
共价催化
酶在催化过程中与底物上的某些基团暂时形成不稳定的共价中间物
亲核试剂
供给一对电子给反应物生成共价键的试剂
亲电试剂
从反应物得到一对电子生成共价键的试剂
通常情况下,路易斯酸都是亲电试剂,路易斯碱都是亲核试剂
底物形变
当酶与底物相遇时,酶诱导底物中的敏感键使其更加敏感,从而产生电子张力发生形变,使得底物更加接近他的过渡态
几种常见酶的结构与功能
丝氨酸蛋白酶
催化基团包括一个不可缺少的丝氨酸残基,DIFP是它的不可逆催化剂
胰蛋白酶
具有很深的底物结合口袋,适合长链Lys和Arg的结合
胰凝乳蛋白酶
底物结合口袋较宽,且口袋壁分布疏水氨基酸,特别适合芳香族氨基酸的结合
弹性蛋白酶
结合口袋浅,适合侧链较小的氨基酸(如甘氨酸)
催化三元体
Ser195
充当进攻底物的亲和基团
共价催化
His57
作为广义的酸碱催化剂
Asp102
定向His57,影响其ph改变其酸碱性质
概要
氧阴离子穴
通过氢键稳定四面体过渡态
丝氨酸蛋白酶催化过程总结
Ser195广义的酸催化,共价催化(进攻底物)
His57广义的酸碱催化
质子转移1
Ser-His His-底物1
质子转移2
H2O-His His-Ser
两次亲核进攻
Ser-O进攻底物羰基,形成共价键(四面体过渡态)
H2O-OH进攻共价复合物羰基,形成共价键(四面体稳定态)
一个氧阴离子穴
通过氢键稳定四面体过渡态
金属蛋白酶
在活性中心结合有金属离子,其活性绝对需要金属离子,因此金属螯合剂EDTA可导致其活性丧失
天冬氨酸蛋白酶
催化基团包括两个天冬氨酸,在偏碱性的ph条件下失活
巯基蛋白酶
催化基团包括一个半胱氨酸残基,碘代乙酸为其不可逆催化剂
酶活性的调节
酶的量变(控制酶水平,浓度)
控制酶基因的表达和酶分子的降解
同工酶
催化相同反应,但结构性质不同的酶
例:己糖激酶的四种同工酶
HK1
主要分布在脑
HK2
主要分布在骨骼肌
HK3
主要分布在白细胞
HK4
主要分布在肝脏(葡萄糖激酶)
组织分布不同,浓度变化不相同,应对机体的不同需求
己糖激酶
受产物反馈抑制
控制糖酵解快慢
葡萄糖激酶
单体酶,不受别构抑制
负责降低血糖,维持稳定性
酶的质变(控制酶活性)
在不改变酶浓度的条件下,对已有酶活性进行调解(低能耗,快速)
别构调节
一些别构酶除活性中心外,还含有别构中心,可以结合一些特殊的配体分子,从而改变酶的构象,导致酶活性发生变化
具有协同效应和不具有协同效应的酶都可以受到别构调节
共价修饰
酶分子在别的酶的催化作用下,发生可逆的共价修饰,这种酶叫做共价修饰调节酶,是对一些代谢起调节作用的关键酶,受激素调节,可以级联放大
有些酶以修饰状态为活性状态
有些酶以脱修饰状态为活性状态
酶共价修饰的几种形式
磷酸化
Tyr,Ser,Thr,His
腺苷酸化
Tyr
尿苷酸化
Tyr
ADP核糖基化
Arg,Gln,Cys,His
甲基化
Glu
共价修饰调节的特点
酶以两种不同修饰或不同活性的形式存在
有共价键的变化
受其他调节因素的影响(如激素)
一般为耗能过程
存在放大效应
水解激活
在酶的催化下,无活性的的酶(酶原)转变为有活性酶的过程
激活机理
在酶的催化下,切去酶原多余的肽段
激活的实质
形成酶的活性中心
生理意义
生物自我保护的一种方法
酶活性的一种调节方式,保证体内代谢正常进行
调节蛋白的激活或抑制
某些蛋白质作为配体与特定的酶结合而调节被结合酶的活性
这种蛋白质称为调节蛋白
激活蛋白
抑制蛋白
酶亚基的聚合与解离
酶原激活的特点
过程伴有酶蛋白一级结构的改变
水解过程,不可逆,一次性的调节
是通过级联效应实现的快速的信号放大过程,以完成特定功能
PS:溶菌酶的催化机理
广义的酸碱催化
Glu35提供质子
静电催化
Asp52(负电荷)稳定带正电荷的过渡态底物
底物形变
糖链为适应酶活性中心的构象,从椅式变为半椅式,使糖苷键更容易发生断裂
米氏常数
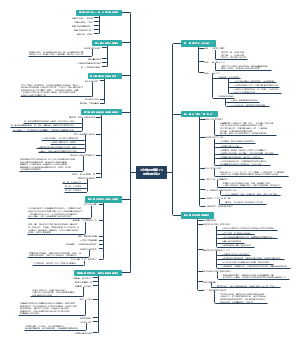
酶
酶的化学本质
酶是由细胞合成的,在机体内行使催化功能的生物催化剂
有酶催化的反应称为酶促反应
底物
在酶催化作用下进行化学反应的物质
产物
酶促反应生成的物质
化学本质
主要是蛋白质,少数是RNA
酶的分类
单纯酶
属于单纯蛋白质的酶
缀合酶
除了蛋白质以外,还结合某些非氨基酸成分(辅助因子)
辅助因子
辅酶
与脱辅酶结合松散,使用透析的方法就可以去除的有机小分子
辅基
以共价键和酶蛋白结合,与脱辅酶结合紧密,不能使用透析或超滤的方法去除的有机小分子
金属离子
酶的催化性质
酶与非酶催化剂的共同性质
只能催化热力学允许的反应
反应完成后本身不被消耗或变化,可以重复使用
对正反应和逆反应的催化作用相同
不改变平衡常数,只加快到平衡的速度或缩短达到平衡的时间
酶特有的催化性质
高效性,专一性
绝对专一性
只作用于一种底物,而不作用于任何其他物质
相对专一性
可作用于一类结构相近的底物
基团专一性
键专一性
立体异构专一性
解释酶专一性三种学说
锁与钥匙学说
诱导契合学说
被广泛认可
三点一线学说
区分对映异构体,解释立体专一
在活性中心与底物结合
活性中心
酶分子上直接与底物结合,并与催化作用直接相关的区域
催化部位
催化能力
结合部位
专一性
多为疏水aa残基,少量亲水aa残基(起催化反应的是亲水aa)
活性中心与底物结构互补,且构象并非固定不变的
反应条件温和,对反应条件敏感,容易失活
具有较高的可调节性
影响酶促反应的因素
酶促反应速率
单位时间内,底物或产物浓度的变化量
mol L/S
影响因素
反应温度
离子强度
质子浓度
影响反应pH
金属离子浓度
可以作为酶的辅因子,从而影响反映速率
反应介质pH
影响酶分子的解离状态
影响底物,辅基或辅酶的解离状态
酶激活剂或酶抑制剂
外因
酶浓度
在一定pH和温度条件下,当底物浓度远远大于酶浓度时,反应速率与酶的浓度呈正比关系
底物浓度
底物浓度的三级反应
底物浓度较低时,呈现一级反应 随着底物浓度的增加,反应表现为混合级反应 底物浓度相当高时,呈现零级反应
酶底物中间物学说
内因
米氏反应动力
描述米氏酶反应速率与底物浓度之间的关系
米氏方程的推导
反应速率为初速率
酶底物复合物处于稳态即ES浓度不发生变化
符合质量作用定律
在稳态时,ES的形成速率与ES的解离速率相等
解读米氏方程
Km是酶反应初速率为Vm的一半是的底物浓度
是酶的特征常数,只与酶的性质有关,与酶的浓度无关
在一定条件下,可以反映酶与底物的亲和力,Km越大,意味着酶与底物的亲和力越低
当酶对底物的Km值小于对产物的Km值,则反应有利于正反应,否则有利于逆反应。
Vmax
理论上的最大反应速率,随着酶浓度的变化而变化
Kcat
酶的催化常数或转换数或周转数,具体指单位时间内,一个酶分子将底物转变为产物的分子总数
kcat=k2=Vmax/Et
可以衡量一个酶促反应的快慢
Kcat/Km
用来衡量酶的催化效率,还可以反应一个酶的完美程度
米氏方程的双重性
当底物浓度很低的时候,即S远小于Km,米氏方程转变为
符合一级动力学
当底物浓度很高的时候,即S远大于Km,米氏方程可转变为
符合零级动力学
米氏酶的双倒数作图
酶抑制剂作用
可逆性抑制剂
以次级键和酶可逆性结合
竞争性抑制剂
抑制剂与底物竞争
非竞争性抑制剂
既能与ES结合,也能与E结合
反竞争性抑制剂
只能与ES结合,使得与活性中心结合的底物将无法再转变为产物
不可逆性抑制剂
以强的化学键与酶不可逆结合,一旦失活不可逆转
基因特异性抑制剂
共价修饰酶活性中心上的必需侧链基团
底物类似物抑制剂
结构上相似于底物,在活性中心与酶结合,不可逆的修饰酶活性中心上的必需基团
过渡态类似物抑制剂
与酶促反应的过渡态极为相似,与酶的活性中心进行结合,导致底物无法与酶结合
自杀型抑制剂
受到酶的催化,发生几步反应,但并不形成产物,专而修饰酶的必需基团导致酶活丧失
别构酶的动力学
别构效应
酶分子的非催化部位与某些化合物可逆的,非共价结合后,发生构象的改变,进而改变酶的活性状态,称为酶的别构效应
具有别够效应的酶称为别构酶
能使酶分子发生别构作用的物质称为别构效应物
别构酶的性质
速率,底物浓度曲线为S型
具有别构效应物
对竞争性抑制表现双向反应
低浓度下,诱发别构酶正协同效应,提高底物的反应速率
高浓度下,占据酶的活性中心,底物与S无法正常结合,抑制反应速率
温和变性可以导致别构效应丧失
通常是寡聚酶,且别构酶占少数
Hill方程
h=1,则不是别构酶,没有底物协同性
双曲线
h>1,速率对底物浓度作图呈S型,具有正底物协同性
S型曲线
酶对环境中底物浓度变化敏感,对代谢进行灵敏的调节
h<1,酶具有负底物协同性
表观双曲线
酶对环境中底物浓度变化极度不敏感,保证重要反应不受底物影响,始终进行下去
酶的催化机制
过渡态稳定学说内容
在任何一个化学反应的过程中,反应物需要达到一个特定的高能状态 才能发生反应,这种不稳定的高能状态被称为过渡态,达到过渡态要求反应物有足够的能量以突破活化能
酶与过渡态的亲和力要远远高于对底物(基态)的亲和力,酶的催化作用来源于对过渡的稳定作用
过渡态稳定的化学机制
邻近定向反应
指两种或两种以上的底物同时在酶的活性中心与酶结合,冻结了某些化学键的平动和转动,从而使其采取正确的空间取向,大大提高了底物的浓度
广义的酸碱催化
指酶的活性中心参与催化的氨基酸残基得到或失去质子(转移到过渡态中间物从而达到稳定过渡态的效果。),这种机制参与绝大多数酶的催化过程
质子供体
质子受体
如果pKa的值接近7,该侧链基团可能是最有效的广义酸碱催化剂,如His残基的咪唑基,在生理条件下,一半为酸,一半为碱,释放质子的速率快
反应速率取决于pH以及缓冲液浓度
静电催化
活性中心电荷的分布用来稳定酶促反应的过渡态,酶使用自身带电基团去中和一个反应过渡态形成时产生的相反电荷而进行催化
有时也会通过酶和底物的静电作用将底物引入酶的活性中心
金属催化
金属酶
含有紧密结合的金属离子
金属激活酶
与溶液中的金属离子松散的结合
金属离子的催化方式
作为路易斯酸发挥作用
与带负电的底物结合,促进底物在反应中正确定向
参与静电催化,稳定带负电的过渡态
通过价态的可逆变化,作为电子供体或电子受体参与氧化还原反应
作为酶结构的组成部分
共价催化
酶在催化过程中与底物上的某些基团暂时形成不稳定的共价中间物
亲核试剂
供给一对电子给反应物生成共价键的试剂
亲电试剂
从反应物得到一对电子生成共价键的试剂
通常情况下,路易斯酸都是亲电试剂,路易斯碱都是亲核试剂
底物形变
当酶与底物相遇时,酶诱导底物中的敏感键使其更加敏感,从而产生电子张力发生形变,使得底物更加接近他的过渡态
几种常见酶的结构与功能
丝氨酸蛋白酶
催化基团包括一个不可缺少的丝氨酸残基,DIFP是它的不可逆催化剂
胰蛋白酶
具有很深的底物结合口袋,适合长链Lys和Arg的结合
胰凝乳蛋白酶
底物结合口袋较宽,且口袋壁分布疏水氨基酸,特别适合芳香族氨基酸的结合
弹性蛋白酶
结合口袋浅,适合侧链较小的氨基酸(如甘氨酸)
催化三元体
Ser195
充当进攻底物的亲和基团
共价催化
His57
作为广义的酸碱催化剂
Asp102
定向His57,影响其ph改变其酸碱性质
概要
氧阴离子穴
通过氢键稳定四面体过渡态
丝氨酸蛋白酶催化过程总结
Ser195广义的酸催化,共价催化(进攻底物)
His57广义的酸碱催化
质子转移1
Ser-His His-底物1
质子转移2
H2O-His His-Ser
两次亲核进攻
Ser-O进攻底物羰基,形成共价键(四面体过渡态)
H2O-OH进攻共价复合物羰基,形成共价键(四面体稳定态)
一个氧阴离子穴
通过氢键稳定四面体过渡态
金属蛋白酶
在活性中心结合有金属离子,其活性绝对需要金属离子,因此金属螯合剂EDTA可导致其活性丧失
天冬氨酸蛋白酶
催化基团包括两个天冬氨酸,在偏碱性的ph条件下失活
巯基蛋白酶
催化基团包括一个半胱氨酸残基,碘代乙酸为其不可逆催化剂
酶活性的调节
酶的量变(控制酶水平,浓度)
控制酶基因的表达和酶分子的降解
同工酶
催化相同反应,但结构性质不同的酶
例:己糖激酶的四种同工酶
HK1
主要分布在脑
HK2
主要分布在骨骼肌
HK3
主要分布在白细胞
HK4
主要分布在肝脏(葡萄糖激酶)
组织分布不同,浓度变化不相同,应对机体的不同需求
己糖激酶
受产物反馈抑制
控制糖酵解快慢
葡萄糖激酶
单体酶,不受别构抑制
负责降低血糖,维持稳定性
酶的质变(控制酶活性)
在不改变酶浓度的条件下,对已有酶活性进行调解(低能耗,快速)
别构调节
一些别构酶除活性中心外,还含有别构中心,可以结合一些特殊的配体分子,从而改变酶的构象,导致酶活性发生变化
具有协同效应和不具有协同效应的酶都可以受到别构调节
共价修饰
酶分子在别的酶的催化作用下,发生可逆的共价修饰,这种酶叫做共价修饰调节酶,是对一些代谢起调节作用的关键酶,受激素调节,可以级联放大
有些酶以修饰状态为活性状态
有些酶以脱修饰状态为活性状态
酶共价修饰的几种形式
磷酸化
Tyr,Ser,Thr,His
腺苷酸化
Tyr
尿苷酸化
Tyr
ADP核糖基化
Arg,Gln,Cys,His
甲基化
Glu
共价修饰调节的特点
酶以两种不同修饰或不同活性的形式存在
有共价键的变化
受其他调节因素的影响(如激素)
一般为耗能过程
存在放大效应
水解激活
在酶的催化下,无活性的的酶(酶原)转变为有活性酶的过程
激活机理
在酶的催化下,切去酶原多余的肽段
激活的实质
形成酶的活性中心
生理意义
生物自我保护的一种方法
酶活性的一种调节方式,保证体内代谢正常进行
调节蛋白的激活或抑制
某些蛋白质作为配体与特定的酶结合而调节被结合酶的活性
这种蛋白质称为调节蛋白
激活蛋白
抑制蛋白
酶亚基的聚合与解离
酶原激活的特点
过程伴有酶蛋白一级结构的改变
水解过程,不可逆,一次性的调节
是通过级联效应实现的快速的信号放大过程,以完成特定功能
PS:溶菌酶的催化机理
广义的酸碱催化
Glu35提供质子
静电催化
Asp52(负电荷)稳定带正电荷的过渡态底物
底物形变
糖链为适应酶活性中心的构象,从椅式变为半椅式,使糖苷键更容易发生断裂
米氏常数