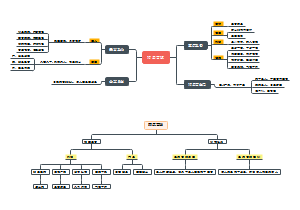
导图社区 微生物 原核生物细胞壁
微生物 原核生物细胞壁
原核微生物细胞壁 原核微生物 回顾 细菌、放线菌、蓝细菌、支、衣、立克次体、螺旋体 多细胞真菌——霉菌单细胞真菌——酵母菌 真菌主要区别:原核细胞 动物和植物 细胞型 是否有核膜包裹的细非细...
编辑于2022-11-28 18:04:20 重庆- 大学专业课资料
他的近期作品
查看更多>>
- 微生物 原核生物细胞壁
原核微生物细胞壁 原核微生物 回顾 细菌、放线菌、蓝细菌、支、衣、立克次体、螺旋体 多细胞真菌——霉菌单细胞真菌——酵母菌 真菌主要区别:原核细胞 动物和植物 细胞型 是否有核膜包裹的细非细...
- 植物生理学 矿质营养
植物生理学矿质营养.ppt,GS、GOGAT、GDH三种酶在细胞内的定位: 1、在绿色组织中:GOGAT存在于叶绿体,GS存在于叶绿体和细胞质,GDH主要存在于线粒体,叶绿体中量很少; 2、在非绿色组织...
- 植物生理学 植物水分生理
①细胞壁以及细胞壁间物质,它们由亲水性的纤维素和果胶质组成。整个植株的细胞壁除根部被凯氏带隔开以外都彼此连接,形成质外体,水分可以在其中自由流动。
微生物 原核生物细胞壁
社区模板帮助中心,点此进入>>
他的近期作品
查看更多>>
- 微生物 原核生物细胞壁
原核微生物细胞壁 原核微生物 回顾 细菌、放线菌、蓝细菌、支、衣、立克次体、螺旋体 多细胞真菌——霉菌单细胞真菌——酵母菌 真菌主要区别:原核细胞 动物和植物 细胞型 是否有核膜包裹的细非细...
- 植物生理学 矿质营养
植物生理学矿质营养.ppt,GS、GOGAT、GDH三种酶在细胞内的定位: 1、在绿色组织中:GOGAT存在于叶绿体,GS存在于叶绿体和细胞质,GDH主要存在于线粒体,叶绿体中量很少; 2、在非绿色组织...
- 植物生理学 植物水分生理
①细胞壁以及细胞壁间物质,它们由亲水性的纤维素和果胶质组成。整个植株的细胞壁除根部被凯氏带隔开以外都彼此连接,形成质外体,水分可以在其中自由流动。
- 相似推荐
- 大纲