导图社区 自私的基因 【英】理查德·道金斯
自私的基因 【英】理查德·道金斯
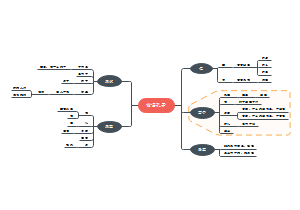
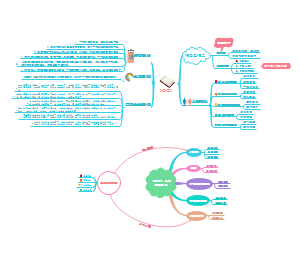
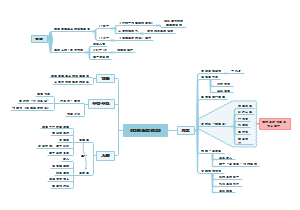
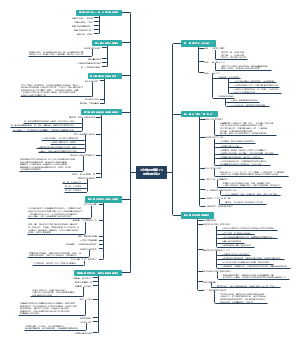
在该书中,道金斯将社会学说中的主要论题逐一做了详细介绍,如利他和利己行为的概念、遗传学上的自私的定义、亲族学说(包括亲子关系和群居昆虫的进化)、性比率学说、相互利他主义、欺骗行为和性差别的自然选择等。同时,道金斯以生物学研究上的进展及自己的理解为基础,将生物进化的单元或层次确定于基因,并通过对伦理学语言的运用,说明基因的基本特性就是“自私”。
编辑于2023-03-28 16:27:03 广东- 自私的基因
- 理查德·道金斯

- 始于极限:女性主义往复书简
在书中,你能感受到两人运用语言的精炼、犀利以及对自己的反思。你能感受到相等阶级的她们,由于时代、父母不同,在“女性主义”、“男人”的态度上也不同。比如“凉美”反复的在追寻一个答案“为何‘鹤子’对男人不感到绝望”;而“鹤子”则好奇“凉美”工作的选择、以及对她的影响......
- 《心智觉醒:我不介意会发生什么》拆书解
你是否也有过感觉被卡住的时候: 也许是不太顺利的职场生涯,让你想去更多地自我实现? 也许是阴霾笼罩的原生家庭,让你很想摆脱父母的束缚? 也许是不堪回首的情感经历,让你渴望彻底走出? 也许是令人纠结的两难选择,让你无法如愿前行? 在这样的环境里,人的心智也会亦步亦趋。怎么办?唯有心智觉醒,提升、拓展、延伸心智的维度,方能破局。 人生破局的底层逻辑是什么? 如何活出自己真正想要的状态? 要从何入手,才能落实到当下? 本书融合东西方哲学、心理学、教练的科学、神经语言程序学、成人发展理论、天赋优势理论等各个学科的理论和方法论,助你从高度、广度和长度三个维度实现心智升维。
- 玩透小红书:素人博主从0到1实战手册
《玩透小红书 ——素人博主从0到1实战手册》是人人可复制的小红书运营指南,主要介绍普通人如何从0开始做小红书博主、如何在小红书上做出既叫好又叫座的内容,从而实现“涨粉”、变现。本书的内容包括账号定位、高效“涨粉”的方法、内容创作、变现、精细化运营、创作心法等,有丰富的案例、实用的工具模板、实操的干货。本书手把手教你从0到1做一个能持续变现的小红书账号。即使你是毫无经验的新手,也能看得懂、拿来即用。
自私的基因 【英】理查德·道金斯
社区模板帮助中心,点此进入>>
- 始于极限:女性主义往复书简
在书中,你能感受到两人运用语言的精炼、犀利以及对自己的反思。你能感受到相等阶级的她们,由于时代、父母不同,在“女性主义”、“男人”的态度上也不同。比如“凉美”反复的在追寻一个答案“为何‘鹤子’对男人不感到绝望”;而“鹤子”则好奇“凉美”工作的选择、以及对她的影响......
- 《心智觉醒:我不介意会发生什么》拆书解
你是否也有过感觉被卡住的时候: 也许是不太顺利的职场生涯,让你想去更多地自我实现? 也许是阴霾笼罩的原生家庭,让你很想摆脱父母的束缚? 也许是不堪回首的情感经历,让你渴望彻底走出? 也许是令人纠结的两难选择,让你无法如愿前行? 在这样的环境里,人的心智也会亦步亦趋。怎么办?唯有心智觉醒,提升、拓展、延伸心智的维度,方能破局。 人生破局的底层逻辑是什么? 如何活出自己真正想要的状态? 要从何入手,才能落实到当下? 本书融合东西方哲学、心理学、教练的科学、神经语言程序学、成人发展理论、天赋优势理论等各个学科的理论和方法论,助你从高度、广度和长度三个维度实现心智升维。
- 玩透小红书:素人博主从0到1实战手册
《玩透小红书 ——素人博主从0到1实战手册》是人人可复制的小红书运营指南,主要介绍普通人如何从0开始做小红书博主、如何在小红书上做出既叫好又叫座的内容,从而实现“涨粉”、变现。本书的内容包括账号定位、高效“涨粉”的方法、内容创作、变现、精细化运营、创作心法等,有丰富的案例、实用的工具模板、实操的干货。本书手把手教你从0到1做一个能持续变现的小红书账号。即使你是毫无经验的新手,也能看得懂、拿来即用。
- 相似推荐
- 大纲