导图社区 四、物质进出细胞
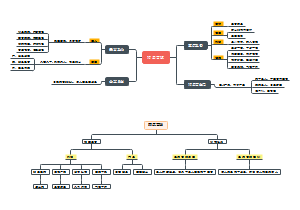
高中生物之物质进出细胞学习笔记,讲述了跨膜运输(被动运输、主动运输)、非跨膜运输(胞吞胞吐)、判断方法(三看法)。
社区模板帮助中心,点此进入>>
《老人与海》思维导图
《钢铁是怎样炼成的》章节概要图
《傅雷家书》思维导图
《阿房宫赋》思维导图
《西游记》思维导图
《水浒传》思维导图
《茶馆》思维导图
《朝花夕拾》篇目思维导图
英语词性
生物必修一
四、物质 进出细胞
方式
跨膜运输
被动运输
自由扩散
运输方向:高浓度→低浓度
特点:不需要能量、不需要载体
实例
水
气体
脂溶性物质
甘油
乙二醇
脂肪酸
胆固醇
尿素
协助扩散
特点
需要载体蛋白,但不需要能量
需要通道蛋白,但不需要能量
载体蛋白协助
红细胞吸收葡萄糖
通道蛋白协助
钠离子进入神经细胞, 钾离子运出神经细胞
主动运输
运输方向:低浓度→高难度
特点:需要能量,需要载体
离子
碘离子,钾离子等等
葡萄糖
氨基酸
非跨膜运输
胞吞胞吐
需要能量,一般不需要载体
依靠细胞膜的流动性
大分子:分泌蛋白
颗粒物质:白细胞的吞噬作用
少数小分子:神经递质
判断方法 (三看法)
一看浓度
高浓度到低浓度
不需要载体
需要载体
低浓度到高浓度
二看能量
不耗能
耗能
根据分子大小
小分子,一般是主动运输
大分子胞吞胞吐
三看物质种类
水,脂溶性物质,气体(自由扩散)
离子,氨基酸,葡萄糖(主动认输)
蛋白质,神经递质(胞吞胞吐)