导图社区 基因表达调控
基因表达调控
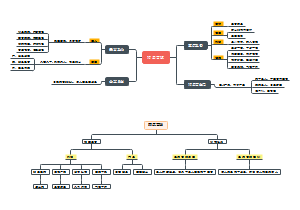
这是一篇关于基因表达调控 的思维导图,生物化学基因表达调控内容,包括真核,原核生物的表达调控,详细介绍了各部分主要内容
提示: 本内容由社区用户上传并分享。平台不对内容的真实性、合法性、知识产权归属及是否侵害第三方权利进行事前审核或保证。本内容可能包含受版权保护的图片、字体或其他第三方素材,使用前请自行确认授权范围。
- 基因表达调控

- 线粒体通透性转换孔(mPTP)组成
线粒体通透性转换(mPT)是一种突然导致低分子量溶质(分子量高达 1,500)穿过通常不可渗透的线粒体内膜的现象。mPT 由线粒体通透性转换孔(mPTP)介导,mPTP 是在内膜和外膜界面组装的超分子实体。mPTP 是位于线粒体内膜上由多种蛋白共同组成的、具有非特异性、电压依赖性的复合物孔道。它的分子组成目前尚不清楚。但普遍认为是由基质的亲环蛋白D(cyclophilin,CyP-D)、内膜的腺嘌呤核苷酸转位酶(adeninenucleotide translocase,ANT)、外膜的电压依赖性离子通道(voltage-dependent anion channel,VDAC)等共同组成。
- PPT制作SOP(汇报的一些逻辑)
组会汇报或者其他汇报,基本上的步骤都是:①罗列背景情景;②当下的冲突有哪些(为什么要做这个工作?有什么工作是别人没解决的?不解决会怎样?)③问题有什么(目前主要做了什么工作,做的过程出现了什么问题?)④分析问题,找出答案
- 了解一个新领域的sop
SOP是Standarded Operating Procedure:标准操作流程,通过把重复性的工作进行细化和拆解,固定成一套流程程序,每天有的放矢,这样就可以拜托低效的忙碌,将主要的时间和精力用在关键任务上,节约时间。
基因表达调控
社区模板帮助中心,点此进入>>
- 线粒体通透性转换孔(mPTP)组成
线粒体通透性转换(mPT)是一种突然导致低分子量溶质(分子量高达 1,500)穿过通常不可渗透的线粒体内膜的现象。mPT 由线粒体通透性转换孔(mPTP)介导,mPTP 是在内膜和外膜界面组装的超分子实体。mPTP 是位于线粒体内膜上由多种蛋白共同组成的、具有非特异性、电压依赖性的复合物孔道。它的分子组成目前尚不清楚。但普遍认为是由基质的亲环蛋白D(cyclophilin,CyP-D)、内膜的腺嘌呤核苷酸转位酶(adeninenucleotide translocase,ANT)、外膜的电压依赖性离子通道(voltage-dependent anion channel,VDAC)等共同组成。
- PPT制作SOP(汇报的一些逻辑)
组会汇报或者其他汇报,基本上的步骤都是:①罗列背景情景;②当下的冲突有哪些(为什么要做这个工作?有什么工作是别人没解决的?不解决会怎样?)③问题有什么(目前主要做了什么工作,做的过程出现了什么问题?)④分析问题,找出答案
- 了解一个新领域的sop
SOP是Standarded Operating Procedure:标准操作流程,通过把重复性的工作进行细化和拆解,固定成一套流程程序,每天有的放矢,这样就可以拜托低效的忙碌,将主要的时间和精力用在关键任务上,节约时间。
- 相似推荐
- 大纲