导图社区 证券从业-金融市场基础知识-第二章-第1节-中国的金融体系
- 64
- 3
- 0
- 举报
证券从业-金融市场基础知识-第二章-第1节-中国的金融体系
证券从业考试-金融市场基础知识-第二章-第1节-中国的金融体系,希望这份脑图会对你有所帮助。
提示: 本内容由社区用户上传并分享。平台不对内容的真实性、合法性、知识产权归属及是否侵害第三方权利进行事前审核或保证。本内容可能包含受版权保护的图片、字体或其他第三方素材,使用前请自行确认授权范围。
- 证券从业
- 中国的金融体系
- 相似推荐
- 大纲
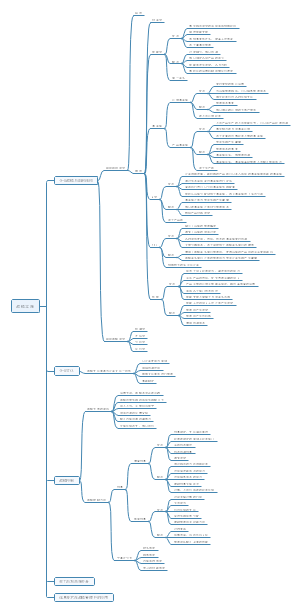
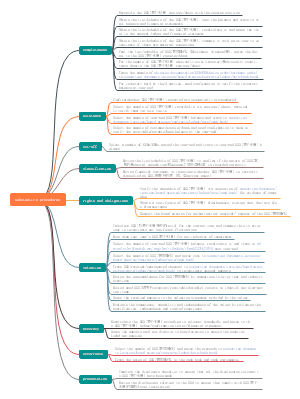
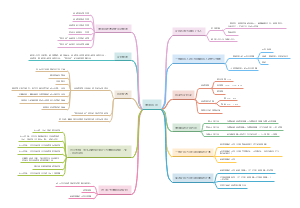
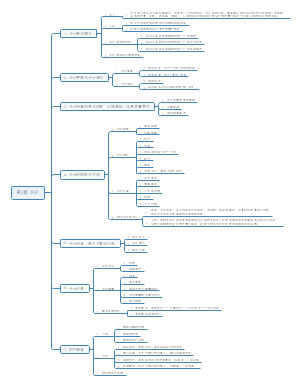
中国的金融体系
建 国 以 来 我 国 金 融 市 场 的 发 展 历 史
( 一)1949~1978年:计划经济时期“大 一 统”的金融体系
1948年12月1日,中国人民银行成立,并于1948年12月7日同时发行人民币。
1. “大”—— 中国人民银行分支机构覆盖全国
2.“ 一 ”—— 中国人民银行是该时期集中央银行和商业银行双重职能于一身唯一的银行
3. “统”—— 全国实行“统存统贷”的信贷资金管理体制
(二)1979~1983年:多元混合型金融体系
1.1979年,恢复设立中国农业银行
2. 中国银行从中国人民银行分设出来
3.中国建设银行从财政部分离出来
4.中国信托投资公司和中国人民保险公司等非银行金融机构先后成立
(三)1984~1993年:以中央银行为中心,国家专业银行为主体,多种金融机构分工协作的金融体系逐步形成
1 . 1984年1月1日起,中国人民银行开始专门行使中央银行职能
2.1985年全面推行“拨改贷”政策
3. 1990年12月,上海证券交易所、深圳证券交易所先后开业,标志着股票市场正式运作的开始
4 . 1992年10月,国务院证券管理委员会和中国证券监督管理委员会成立,标志着中国资本市场开始逐步纳入全国统一监管框架。
(四)1994~2003年:逐步改革和完善的市场化金融体系
1.成立了国家开发银行、中国进出口银行、中国农业发展银行政策性银行;
2.成立信达、华融、长城、东方四家资产管理公司;
3.银行业与信托业、证券业分离,银行不得从事证券业经营;
4.建立以市场为基础的、有管理的浮动汇率制度和统一规范的外汇市场。
(五)2004~2011年:多种所有制和多种经营形式、结构合理、功能完善、高效安全的现代金融体系
2003年4月,中国银行业监督管理委员会成立,标志着我国“分业经营、分业监管”的金融体系得以确立。
(六)2012年至今:基本金融制度逐步健全、对外开放程度不断提高的现代金融体系
我国金融市场的发展现状
一、货币市场(1984年银行间同业拆借市场建立)
(一)银行间同业拆借市场
(二)短期债券市场
(三)债券回购市场
(四)票据贴现市场
二、 债券市场
(一)场外市场
1.银行间市场(场外债券批发市场)
(1)参与者限定为机构
(2)中央国债登记公司托管
2.银行柜台市场(场外债券零售市场)
(1)参与者限定为个人
(2)中央国债登记结算公司一级托管,商业银行二级托管
(二)场内市场
1.上海证券交易所
中国证券登记结算公司上海分公司托管
2.深圳证券交易所
中国证券登记结算公司深圳分公司托管
(三)2020年,我国债券市场运行特点
1.大力发行疫情防控债券
2.债券市场产品创新提速
3.债券市场制度建设持续推进
4.债券市场对外开放全面推进
三、 股票市场
( 一 )重要事件
1.2005年完成股权分置改革
2.截至2020年末,沪、深两地上市公司数量达到4233家,总市值达79.72万亿元。
( 二 ) 分 类
1.场内市场
(1)沪深主板市场
(2)中小板市场
(3)科创板市场
(4)创业板市场
(5)全国中小企业股份转让系统(新三板)
2.场外市场
(1)区域性股权交易市场
(2)交易柜台市场
四、 外汇市场
(一)1994年,建立了全国统一 的银行间外汇市场,实现了人民币官方汇率与外汇调剂市场汇率并轨。
(二)2005年7月21日,人民币汇率形成机制改革后,外汇市场发展迅速,交易方式、时间、品种、清算等各个方面不断丰富完善。
五、 黄金市场
(一)2002年10月,上海黄金交易所正式运行,标志着我国国内黄金市场的正式开放。
(二)中国黄金市场目前初步形成了以上海黄金交易所黄金现货为主体,上海期货交易所黄金期货、商业银行柜台黄金产品共同组成的,层次分明且各有侧重的多元市场体系。
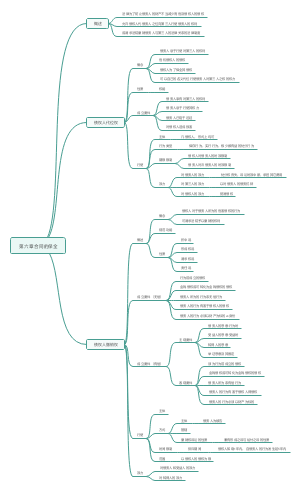
六、 商品期货市场
( 一 )交易所
1.1990年10月我国成立了第一家商品期货交易所—— 郑州商品交易所
2. 目前,我国成立了上海期货交易所、大连商品交易所、郑州商品交易所和广州期货交易所四大商品期货交易所。
3.2021年4月19日,广州期货交易所揭牌仪式举行。
(二)监管(五位 一 体)
1.中国证监会
2.地方证监局
3.期货交易所
4.期货市场监控中心
5 . 中国期货业协会
七 、 金融衍生品市场
( 一 ) 大 事 记
1 . 2006年9月,中国金融期货交易所正式成立
2 . 2 0 1 0 年 4 月 1 6 日 , 中 国 金 融 期 货 交 易 所 推 出 沪 深 3 0 0 股 指 期 货 合 约
3 . 2 0 1 5 年 2 月 9 日 , 我 国 首 个 场 内 金 融 期 权 — — 上 证 5 0ETF 期 权 , 在上海证券交易所正式上市交 易
( 二 ) 交 易 品 种
1 .利率类衍生品
2 .权益类衍生品
3.货币类衍生品
4.信用类衍生品
影响我国金融市场运行的主要因素
一 、经济因素
( 一)宏观经济运行
1.国内生产总值 (GDP)
2.经济周期——复苏、繁荣、衰退、萧条
3.通货膨胀与利率
4.汇率和国际资本流动
(二)宏观经济政策
1.货币政策
2.财政政策
(1)税收政策
(2)公共支出政策
(3)国债发行
3.收入政策
(1)收入总量目标
着眼于近期的宏观经济总量平衡,着重处理积累和消费、人们近期生活水平改善和国家长远经济发展的关系以 及失业和通货膨胀的问题。
(2)收入结构目标
着眼于处理各种收入的比例,以解决公共消费和私人消费、收入差距等问题。
(三)国际经济环境因素
1.经济全球化
2.放松管制和加强管制两种经济哲学的交替
3.世界货币制度
二、 法律因素
完备的金融法律和高效的执法效率是金融市场平稳运行的根本保障。
三、 市场因素
四 、技 术 因 素
广泛应用促使全球统一金融市场的形成,国际金融与国内金融的界限日益模糊。
五 、心 理 因 素
六 、体 制 或 管 理 因 素
七 、 其 他 因 素
( 一 ) 文 化 因 素
( 二 ) 人 口 因 素
金 融 中 介 机 构 体 系 的 构 成
一 、金 融 中 介 机 构 的 定 义
( 一 ) 狭 义 的 金 融 机 构
在金融活动中,处于信用关系中借贷双方之间的中介,从贷方那里借入资金,再向借方贷出,其分别与借方和贷方形成独立的债权和债务关系, 一般指商业银行。
( 二 ) 广 义 的 金 融 中 介 机 构
泛指参与或服务于金融市场活动而获取收益的各类组织和机构。
二 、金 融 中 介 机 构 的 分 类
1.金融中介机构的分类
按业务特征及货币创造、交换媒介和支付手段的能力划分
银行金融中介机构
非银行金融中介机构
按是否承担国家政策性金融业务
政策性金融机构
商业性金融机构
按所从事业务的性质及其在金融活 动中所起的作用
投资类金融中介机构
保障类金融中介机构
服务类金融中介机构
2.按经济活动金融中介机构的分类
不包含保险和养老基金的 金融中介活动
货币中介;中央银行和其他货币中介(主要是指存款货币银行)的活动;其他金融中介;金融租赁以及其他如农业信贷、进出口信贷、消费信贷等专业信贷的活动
保险和养老基金(不包括强 制性社会保障)
生命保险活动;养老基金活动;非生命保险活动
辅助金融中介的活动
投资银行、投资基金之类的证券交易活动;与金融中介有关的其他辅助活动
3.国民核算体系 SNA 按机构的分类
(1)中央银行
(2)其他存款公司
(3)不是通过吸纳存款筹集资金的投资公司、金融租赁公司,以及消费信贷公司
(4)金融辅助机构
(5)保险公司和养老基金
三 、我国的金融中介机构体系
( 一)以中央银行为主导、国有商业银行为主体的多层次银行机构体系
(二)以证券公司、期货公司和证券投资基金为主,以各类投资咨询中介、信托机构为辅的多元化投资中介体系
(三)人寿保险公司、财产保险公司、再保险公司以及提供多种多样保险服务的保险中介体系。
商业银行、证券公司、保险公司等主要金融中介机构的业务
一 、商业银行的业务
资产负债表是商业银行业务活动的总括反应。
负债业务
◆ 商业银行组织资金来源的业务活动;
商业银行的负债包括自有资金和吸引外来资金两大部分。
①自有资金:(股本金、储备资金、未分配利润
②存款负债70%以上
③借款负债:中央银行借款、同业借款、回购协议、国外市场借款
资产业务
将其吸收的资金贷放或投资出去赚取收益的活动;
主要包括现金资产、贷款业务、票据贴现、投资业务等。
表外业务
◆ 商业银行从事的,按照现行的会计准则不计入资产负债表内,不形成现实资产负债,但能够引 起当期损益变动的业务。
◆ 担保承诺类、代理投融资服务类、中介服务类、其他类等
中间业务
◆ 不构成商业银行表内资产、表内负债,形成银行非利息收入
银行利用自身在机构网点、技术手段和信息处理等方面的优势,代理客户承办收付和委托事项, 并收取手续费的业务
二 、证券公司的业务
( 一 )证券经纪业务
(二)证券投资咨询业务
(三)与证券交易、证券投资活动有关的财务顾问业务
(四)证券承销与保荐业务
(五)证券自营业务
(六)证券资产管理业务
( 七 ) 其 他 业 务
1.融资融券业务
2.证券公司中间介绍业务
3.私募投资基金业务
4.另类投资业务等
三 、保险公司的业务
财产保险公司
基础类业务
机动车保险;企业/家庭财产保险及工程保险;责 任保险;船舶/货运保险;短期健康/意外伤害保 险 。
扩展类业务
农业保险;特殊风险保险;信用 保证保险;投资型保险
人身保险公司
基础类业务
普通型保险;健康保险;意外伤害保险;分红型保险;万能型保险。
扩展类业务
投资连结型保险;变额年金。
我国银行业、证券业、保险业、信托业的行业基本情况
一 、我国银行业的基本情况
( 一 )4家特殊金融机构(政策性银行等)
( 二 ) 7 8 家 银 行
(三)133家城市商业银行
(四)3898家农村金融机构
(五)123家银行附属机构(金融租赁公司等)
(六)5家金融资产管理公司、5家货币经纪公司、68家信托公司、31家其他金融机构
(七)256家企业集团财务公司
二 、我国证券业的基本情况
加快财富管理业务转型,服务市场投资理财需求。
三 、我 国 保 险 业 的 基 本 情 况 ( 新 修 正 )
截至2020年底,我国共有各类保险机构233家。
四 、我国信托业的基本情况
截至2020年底,全国共有68家信托公司
我 国 金 融 市 场 的 监 管 架 构
一 、我国金融监管体制的演变
( 一 ) 体 制 变 革
1.变革前——“ 一行三会”
(1)中国人民银行
(2)证监会
(3)银监会
(4)保监会
2.变革后“ 一委一行两会”
(1)国务院金融稳定发展委员会
(2)中国人民银行
(3)证监会
(4)中国银行保险监督管理委员会
( 二 ) 演 变 阶 段
1.第一阶段:1992年以前—— 中国人民银行集中统一监管
2.第二阶段:1992年到1997年— 一 国务院证券委员会和中国证券监督管理委员会成立,国务院证券委员会负责指导、监督和归口管理中国证监会。
3.第三阶段:1998年到2008年
4.第四阶段:2009年到2016年——“ 一行三会”分业监管的金融监管体制正式确立金融监管体制开始了金融监管的协调性和有效性的初步改革
5.第五阶段:2017年以来—— 金融监管体制进入“ 一委一行两会”的新阶段
“ 一 委 一 行 两 会 ” 的 职 责
一 、国务院金融稳定发展委员会的职责
( 一 ) 落 实 党 中 央 、 国 务 院 关 于 金 融 工 作 的 决 策 部 署 ;
( 二 ) 审 议 金 融 业 改 革 发 展 重 大 规 划 ;
(三)统筹金融改革发展与监管,协调货币政策与金融监管相关事项,统筹协调金融监管重大事项,协调金融政策与相关财政政策、产业政策等;
(四)分析研判国际国内金融形势,做好国际金融风险应对,研究系统性金融风险防范处置和维护金融稳定重大政策;
(五)指导地方金融改革发展与监管,对金融管理部门和地方政府进行业务监督和履职问责等。
二 、中 国 人 民 银 行 的 职 责
制定和执行货币政策,维护金融稳定,承担一定的金融服务。
◆ 拟订金融业改革、开放和发展规划,承担综合研究并协调解决金融运行中的重大问题、促进金融业协调健康发展的责任。
◆ 牵头建立宏观审慎管理框架, 拟订金融业重大法律法规和其他有关法律法规草案,制定审慎监管基本制 度,建立健全金融消费者保护基本制度。
◆ 监督管理银行间债券市场、货币市场、外汇市场、票据市场、黄金市场及与上述市场有关的场外衍生产品。
◆ 牵头负责重要金融基础设施建设规划并统筹实施监管,推进金融基础设施改革与互联互通,统筹互联网金融监管工作。
三 、中 国 银 行 保 险 监 督 管 理 委 员 会 的 职 责
参考《中国银行保险监督管理委员会职能配置、内设机构和人员编制规定》
四 、中 国 证 券 监 督 管 理 委 员 会 的 职 责
参考《中国证券监督管理委员会职能配置、内设机构和人员编制规定》
金融服务实体经济的要求
一、有关金融与实体经济关系的理论与国际经验
二、金融危机的教训与宏观审慎监管的兴起
三、我国增强金融服务实体经济能力的障碍与挑战
(一)所谓金融服务实体经济,实质就是有效发挥其媒介资源配置的功能
(二)所谓为实体经济提供更好地金融服务,就是要求降低流通成本,提高金融的中介效率和分 配效率。
四、经济高质量发展阶段如何增强金融服务实体经济的能力
(一)守住不发生系统性金融风险的底线
(二)健全多层次资本市场体系,补齐各类金融市场的短板
(三)深化金融监管体制改革,加强监管协同,补齐监管空白
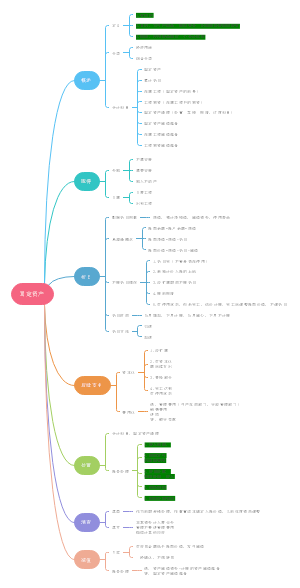
中央银行的业务(资产负债表)和主要职能
一、中央银行的主要职能
(一)中国人民银行的主要职能
1.制定和执行货币政策
2 .防范和化解金融风险
3 .维护金融稳定
(二)中央银行职能的概括
发行的银行
是全国唯一有权发行货币的机构;
银行的银行
中央银行与商业银行之间带有管理和被管理性质的特殊关系;
只与商业银行和其他金融机构发生业务往来,并不与企业和个人发生直接的信用关系;
业务:集中保管商业银行的存款准备金、作为商业银行的最后贷款人、组织全国商业银行之间的 清算。
政府的银行
代表国家制定和执行货币政策,处理对外金融关系,为国家提供各种金融服务。
中央银行作为“政府的银行”,其具体职能包括:
1.代理国库收支;
2 .代理政府债券发行;
3 .为政府提供信贷;
4.代表政府管理金融活动,制定、发布并监督执行有关金融和业务的命令及规章,负责金融业的统 计、调查、分析和预测;
5.持有、管理、经营国家外汇储备和黄金储备;
6.代表政府参加国际金融组织活动,进行国际金融合作。
二、中央银行的业务(资产负债表)
资产方
国外资产
外汇
货币黄金
其他国外资产
对政府债权
对其他存款性公司债权
对其他金融性公司债权
对非金融性部门债权
其他资产
总资产
负债方
储备货币
货币发行
其他存款性公司存款
不计入储备货币的金融性公司存款
发行债券
政府存款
自有资金
其他负债
总负债
存款准备金制度与货币乘数的概念
一、存款准备金制度
(一)概念
国家以法律形式规定存款性金融机构所吸收的存款必须按一定比例,向中央银行缴纳一定准备金的制度。
(二)分类
1.法定存款准备金率一一以法律的形式确定的商业银行缴存中央银行的存款占其吸收存款的比例。法 定准备率越高,则银行吸收的存款用于贷款的数额就越少,货币供应量也会越少;反之,则越多。
2 .超额准备金一一商业银行准备金中超过法定的部分
二、货币乘数
(—)概念
基础货币和货币供给之间的倍数关系,被称为货币乘数。
(二)计算
1.简单货币乘数
D1 = D0 x 1/r
D0为初始存款,r为法定存款准备金率
D0和Di之间存在乘数关系,即为货币乘数。若以m代表货币乘数,则公式为:m =1/r
2.完整的货币乘数公式:m =c +1/c + r + e;C为提现率(客户提取的现金与活期存款总额之比),e为超额准备金率
货币政策的概念及目标
一、货币政策的概念
中央银行利用其掌握的利率、汇率、借贷、货币发行、外汇管理及金融法规等工具,采用的各种控制和调节货 币供给量或信贷规模的方针、政策和措施的总称。
二、货币政策的目标
操作目标
中介目标
最终目标
•短期货币市场利率
货币供应量
稳定物价(首要)
•基础货币
长期利率
充分就业
银行准备金
银行信贷规模
促进经济增长
1.货币政策的目标
中介目标又被称为远期目标,是货币政策作用过程中一个十分重要的中间环节,也是判断货币政策力度和效果的重要指示变量。
利率
优点:可控性强、可測性强、相关性强。
缺点:抗干扰性较差,容易受心理预期、金融市场投机活动等各种非货币政策因素的影响,从 而降低其真实性。
货币供应量
优点:可測性、可控性和相关性似乎都要比利率指标略胜一筹。
缺点:但随着金融创新和金融管制的放松,货币供应量日益成为模糊的、难以控制的指标。
2.目标选取的标准
可測性一一信息易获得并能量化
可控性一一能被中央银行的各种货币政策工具有效地控制和调节
相关性一一与货币政策最终目标之间具有高度相关性
抗干扰性一一中介目标受非政策性因素的影响较小
适应性一一与特定的经济、金融体制有较好的适应性
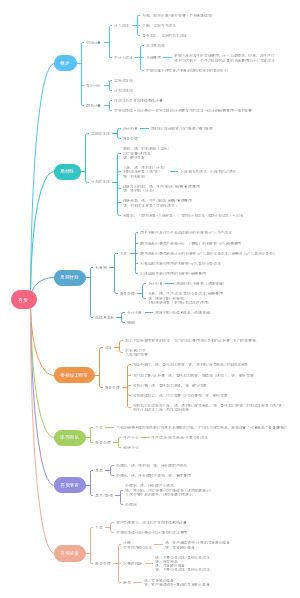
货币政策工具的概念及作用原理
一 、一 般性货币政策工具
存款准备金制度
中央银行通过调整法定存款准备金率来影响商业银行信贷规模,从而间接地对社会货币供应量进行控制的政策措施。
再贴现政策
中央银行(处于被动地位)通过直接调整或制定对合格票据的贴现利率,来干预和影响市场利 率,从而调节货币供应量的政策措施。
公开市场业务
中央银行在证券市场上公开买卖各种有价证券。
债券交易主要包括回购交易、现券交易和发行中央银行票据。
正回购到期和逆回购为央行向市场投放流动性的操作。
正回购和逆回购到期为央行向市场收回流动性的操作。
主动权在央行,可以进行经常性、连续性的操作,有较强的可逆转性。
二、 选择性货币政策工具
(一)消费者信用控制(如信用卡分期付款最长分期次数)
(二)证券市场信用控制(如两融业务最低保证金比例)
(三)不动产信用控制(如首套房首付30%)
(四)直接信用控制(如规定的不同期限存款利率)
(五)间接信用控制
1.道义劝告(如要求配合央行货币政策实施)
2.窗口指导(如规定银行季度贷款增减额——非强制)
三、 创新型货币政策工具
( 一)短期流动性调节工具 (SLO)
1.银行体系流动性出现临时性波动时使用
2.7天期内短期回购为主
(二) 常备借贷便利 (SLF)
1.满足金融机构期限较长的大额流动性需求
2. 以隔夜和7天为主,最长期限为3个月
(三)抵押补充贷款 (PSL)
1.支持经济重点领域和薄弱环节
2.操作对象主要为政策性银行
(四)中期借贷便利 (MLF)
1.对象为符合宏观审慎管理要求的商业银行、政策性银行
2.采取质押方式发放,并需提供国债、央行票据、政策性金融债、高等级信用债等优质债券作为合格质押品
(五)临时流动性便利 (TLF)
向现金投放中占比高的几家大型商业银行提供临时性流动性支持,以满足春节前后现金投放的集中需求。
(六)临时准备金动用安排 (CRA)
允许在现金投放中占比较高的全国性商业银行在春节期间出现临时流动性缺口时,可临时使用不超过2个百分点的法定存款准备金,使用期限为30天。
(七)定向中期借贷便利 (TM LF)
定向支持金融机构向小微企业和民营企业发放贷款。
货 币 政 策 的 传 导 机 制
一 、传导机制的理论
操作目标一中介目标一最终目标
二 、传导机制分类
( 一 )利率传导机制
货币供应量个→实际利率↓→投资个→总产出个
(二)信用传导机制
1.银行信贷渠道的运行机制:
货币供应量个→银行存款和贷款个→投资个→总产出个
2.企业:
货币供应量个→股价个→净值个→逆向选择和道德风险↓→贷款个→投资个→总产出个
( 三 ) 资 产 价 格 传 导 机 制
1.托宾q 理 论
货币供应量个 →股价个 →托宾q 个→投资个→总产出个
托宾q 值为企业的市场价值与资本重置成本之比
2.莫迪利安尼理论:
货币供应量个→股价个→金融资产价值个→财务困难的可能性↓→耐用消费品和住宅支出个→总产出个
(四)汇率传导机制
货币供应量个一实际利率↓ → 汇率↓ →净出口个 → 总产出个
我 国 金 融 业 进 一 步 对 外 开 放 的 背 景 和 主 要 政 策 措 施
一、背景
( 一 ) 金 融 业 对 外 开 放 是 我 国 加 入WTO 的 承 诺
( 二 ) 金 融 业 对 外 开 放 是 我 国 实 现 高 质 量 发 展 的 内 在 要 求
( 三 ) 金 融 业 开 放 是 中 国 深 度 融 入 全 球 经 济 的 必 然 选 择
(四)扩大金融业对外开放有利于构建多元化的金融体系,推动金融供给侧结构性改革
二 、主 要 政 策 措 施
人大 国家主席 博鳌亚洲论坛 2018年4月10日 大幅度放开金融行业 准入
央行 央行行长 博鳌亚洲论坛分论坛 2018年4月11日 3大原则、12大措施
银行保险业监管
银保监会
2018年4月27日 4个方面15条开放 措施
2019年5月01日 12条进一步开放措 施
三 、措施内容
( 一 )央行的三大原则
1.准入前国民待遇和负面清单原则
2.对外开放将与汇率形成机制改革和资本项目可兑换进程相互配合,共同推进
3.开放的同时要重视防范金融风险,要使金融监管能力与金融开放度相匹配
(二)央行的十二大措施在证监会、银保监会措施中的细化和落地
涉及银行业的第一条
取消银行和金融资产管理公司的外资持股比例限制,内外资一视同仁;允许外国银行在我国境内同时设立分行和子行
银保监会措施
1.取消对中资银行和金融资产管理公司的外资持股比例限制,实施内外一致的股权投资比例规则
2.放宽外资设立机构条件,包括允许外国银行在中国境内同时设有子行和分行
3.对外国银行境内分行实施合并考核,调整外国银行分行营运资金管理要求
4.放宽中外合资银行中方股东限制,取消中方唯一或主要股东必须是金融机构的要求
5.取消单家中资银行和单家外资银行对中资商业银行的持股比例上限
6.取消外国银行来华设立外资法人银行的100亿美元总资产要求和外国银行来华设立分行的200亿美元总资产要求
涉及银行业的第二条
鼓励在信托、金融租赁、汽车金融、货币经纪、消费金融等银行业金融领域引入外资
银保监会措施
1.鼓励信托、金融租赁、汽车金融、货币经纪、消费金融等各类银行业金融机构引进境外专业投资者
2.取消境外金融机构投资入股信托公司的10亿美元总资产要求
3.鼓励和支持境外金融机构与民营资本控股的银行业机构开展股权、业务和技术等各类合作
4.按照内外资一致原则,放宽中资和外资金融机构投资设立消费金融公司方面的准入政策
涉及银行业的第三条
对商业银行新发起设立的金融资产投资公司和理财公司的外资持股比例不设上限银保监会措施
对商业银行新发起设立的金融资产投资公司和理财公司,外资持股比例不设限制
涉及银行业的第四条
大幅度扩大外资银行业务范围
银保监会措施
1 . 全面取消外资银行申请人民币业务需满足开业1年的等待期要求,允许外国银行分行从事“代理发行、代理兑付、承销政府债券”业务,降低外国银行分行吸收单笔人民币定期零售存款的门槛至50万元 ;
2 . 取消外资银行开办人民币业务审批,允许外资银行开业时即可经营人民币业务;
3 . 允许外资银行经营”代理收付款项"业务;
涉及保险业的第一条
允许符合条件的外国投资者来华经营保险代理业务和保险公估业务
银保监会的措施
1 . 允许符合条件的境外投资者来华经营保险代理业务和保险公估业务
2 . 允许境外金融机构入股在华外资保险公司
3 . 允许外国保险集团公司投资设立保险类机构
涉及保险业的第二条
放开外资保险经纪公司经营范围,与中资机构一致
银保监会的措施
1 . 允许境内外资保险集团公司参照中资保险集团公司资质要求发起设立保险类机构
2 .鼓励和支持境外金融机构与民营资本控股的保险机构开展股权、业务和技术等各类合作
涉及保险业的第三条
全面取消外资保险公司设立前需开设2年代表处要求
银保监会的措施
1 . 在全国范围内取消外资保险机构设立前需开设2年代表处要求
2 . 取消外国保险经纪公司在华经营保险经纪业务需满足30年经营年限、总资产不少于2亿美元的要求
涉及保险业的第四条
将保险公司外资持股比例上限放宽至51%,三年后不再设限
银保监会的措施
将外资人身险公司外方持股比例放宽至51%,3年后不再设限
涉及证券业的第一条
将证券公司、基金管理公司、期货公司的外资持股比例上限放宽至51%,三年后不再设限
证监会的措施
按内外资一致原则,允许合资证券和基金管理公司的境外股东实现"一参一控"
涉及证券业的第二条
不再要求合资证券公司境内股东至少有一家是证券公司
证监会的措施
无
涉及证券业的第三条
进一步完善内地与香港两地股票市场互联互通机制,互联互通每日额度扩大4倍
证监会的措施
无
涉及证券业的第四条
不再对合资证券公司业务范围单独设限,内外资一致
证监会的措施
放开外资私募证券投资基金管理人管理的私募产品参与“沪港通”深港通”交易的限制
涉及证券业的第五条
经过中英双方共同努力,沪伦通的准备工作进展顺利,在2018年年内开通
证监会的措施
无
证监会的其他措施
1.推动修订QFⅡ 和 RQFⅡI 制度规则, 进一步便利境外机构投资者参与中国资本市场
2.合理设置综合类券商控股股东的资质要求,特别是净资产要求
3.全面推开日股“全流通”改革
4.适当考虑外资银行母行资产规模和业务经验,放宽外资银行在华从事证券投资基金托管业务的准入限 制
5.持续加大期货市场开放力度,扩大特定品种范围
6.研究扩大交易所债券市场对外开放,拓展境外机构投资者进入交易所债券市场的渠道
7.研究制定交易所熊猫债管理办法,更加便利境外机构发债融资
金稳委的补充措施
1.允许外资机构在华开展信用评级业务时,可以对银行间债券市场和交易所债券市场的所有种类债券评级。
2.鼓励境外金融机构参与设立、投资入股商业银行理财子公司。
3.允许境外资产管理机构与中资银行或保险公司的子公司合资设立由外方控股的理财公司。
4.允许境外金融机构投资设立、参股养老金管理公司。
5.支持外资全资设立或参股货币经纪公司。
6.人身险外资股占比限制从51%提高至100%的过渡期,由原定2021年提前到2020年。
7 . 取 消 境 内 保 险 公 司 合 计 持 有 保 险 资 产 管 理 公 司 的 股 份 不 得 低 于 7 5 % 的 规 定 , 允 许 境 外 投 资 者 持 有 股份 超 过 2 5 % 。