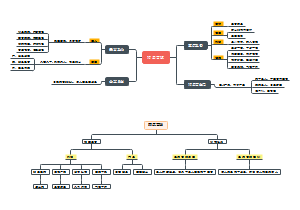
导图社区 RNA的合成
RNA的合成
核糖核酸(缩写为RNA,即Ribonucleic Acid),存在于生物细胞以及部分病毒、类病毒中的遗传信息载体。RNA由核糖核苷酸经磷酸二酯键缩合而成长链状分子。一个核糖核苷酸分子由磷酸,核糖和碱基构成。RNA的碱基主要有4种,即A(腺嘌呤)、G(鸟嘌呤)、C(胞嘧啶)、U(尿嘧啶),其中,U(尿嘧啶)取代了DNA中的T(胸腺嘧啶)。核糖核酸在体内的作用主要是引导蛋白质的合成
提示: 本内容由社区用户上传并分享。平台不对内容的真实性、合法性、知识产权归属及是否侵害第三方权利进行事前审核或保证。本内容可能包含受版权保护的图片、字体或其他第三方素材,使用前请自行确认授权范围。
- RNA的合成
- 诊断小结
这是一篇关于诊断小结的思维导图,主要内容包括:真性血尿的确认,病史询问与体格检查,尿红细胞形态学检查,基础实验室检查,影像学检查,病因分类,对因治疗,不明原因血尿的随访复查。
- 蛋白质的合成
总结了蛋白质的合成的第一节、蛋白质合成体系;第二节、氨基酸与tRNA的连接;第三节、肽链的合成过程等内容,方便大家备考时翻阅查看,提高复习效率。
- RNA的合成
核糖核酸(缩写为RNA,即Ribonucleic Acid),存在于生物细胞以及部分病毒、类病毒中的遗传信息载体。RNA由核糖核苷酸经磷酸二酯键缩合而成长链状分子。一个核糖核苷酸分子由磷酸,核糖和碱基构成。RNA的碱基主要有4种,即A(腺嘌呤)、G(鸟嘌呤)、C(胞嘧啶)、U(尿嘧啶),其中,U(尿嘧啶)取代了DNA中的T(胸腺嘧啶)。核糖核酸在体内的作用主要是引导蛋白质的合成
RNA的合成
社区模板帮助中心,点此进入>>
- 诊断小结
这是一篇关于诊断小结的思维导图,主要内容包括:真性血尿的确认,病史询问与体格检查,尿红细胞形态学检查,基础实验室检查,影像学检查,病因分类,对因治疗,不明原因血尿的随访复查。
- 蛋白质的合成
总结了蛋白质的合成的第一节、蛋白质合成体系;第二节、氨基酸与tRNA的连接;第三节、肽链的合成过程等内容,方便大家备考时翻阅查看,提高复习效率。
- RNA的合成
核糖核酸(缩写为RNA,即Ribonucleic Acid),存在于生物细胞以及部分病毒、类病毒中的遗传信息载体。RNA由核糖核苷酸经磷酸二酯键缩合而成长链状分子。一个核糖核苷酸分子由磷酸,核糖和碱基构成。RNA的碱基主要有4种,即A(腺嘌呤)、G(鸟嘌呤)、C(胞嘧啶)、U(尿嘧啶),其中,U(尿嘧啶)取代了DNA中的T(胸腺嘧啶)。核糖核酸在体内的作用主要是引导蛋白质的合成
- 相似推荐
- 大纲