导图社区 原核生物转录及调控
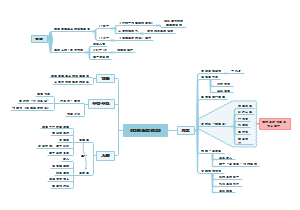
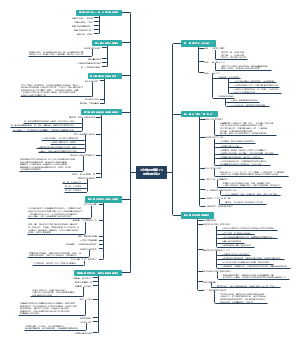
下图整理了原核生物转录及调控,知识点有原核生物的转录、原核生物的转录调控两大板块,适用于考试复习的同学们!
社区模板帮助中心,点此进入>>
论语孔子简单思维导图
《傅雷家书》思维导图
《童年》读书笔记
《茶馆》思维导图
《朝花夕拾》篇目思维导图
《昆虫记》思维导图
《安徒生童话》思维导图
《鲁滨逊漂流记》读书笔记
《这样读书就够了》读书笔记
妈妈必读:一张0-1岁孩子认知发展的精确时间表
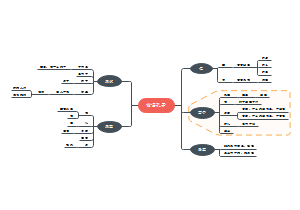
原核生物的转录及其调控
原核生物的转录
基本原则
转录概述
以DNA为模板合成RNA的过程,由RNA聚合酶催化ATP、GTP、UTP、CTP4种原料沿5'到3'方向合成
模板链【template strand】
DNA上通常只有一条链会转录成RNA,也叫反义链【Antisense strand】
编码链【coding strand】
另一条链与RNA序列相同,为有义链【sense strand】
起始
形成闭合复合物
转变为开口复合物
DNA融化:-11——+3
聚合酶结构改变
起始需要起始NTP(常为A)置入活性位点
流产式起始
需要将δ3.2区域逐出RNA出口通道,酶要多次尝试
起始调节
activator
RNA Plo结合后帮助其结合
机制
协同结合
变构
DNA弯曲,远距离作用
repressor
与DNA结合区域与启动子重叠部分,阻碍Pol结合
阻止形成开放复合物
锁定启动子使停留在流产式起始
可变δ因子
不同的亚基结合相同的Rna聚合酶,却结合启动子不同位点
大肠杆菌当温度高时,热休克;δ32耐高温
延伸
聚合酶沿5’--3‘方向延伸新链
DNA进入蟹钳;分叉链分离;NTP加入;后方DNA退火
起始后调控:色氨酸操纵子
终止
发生在终止子【terminator】处,该序列含有自身互补区,能在RNA产物上形成茎环或发夹结构从而停止转录
不依赖Rho的(强)
短的反向重复序列形成发卡结构
一段富含U的序列(使释放)
依赖Rho的(弱)
Rho有ATPase和解旋酶活性
与RNA产物结合,使Pol、Rho蛋白构象改变,易脱离
大肠杆菌RNA聚合酶
只有一种:核心酶(核心酶+δ->全酶)
结构特征
两个亚基---蟹钳结构
DNA从其间进入
催化分叉
链分离
校正功能
焦磷酸键解:错误核酸+ppi->去除一个错误插入的
水解
向后追踪几个并移除
E.coli的δ70因子
结构
+1:翻译起始
两个保守序列:-10,-35(共有序列),接近这些的启动子通常更强(一定时间转录起始量)
上游元件:可提供Rna Pol更多相互位点,提高其结合
extended -10:存在于无-35的启动子上,补偿缺失
因子
αCTD识别上游元件
区域2:识别-10
区域3:识别extended-10
区域4:识别-35
螺旋转角螺旋
δ3.2:在RNA出口通道中间
原核生物的转录调控
概述
操纵子【operon】:由结构基因、 调控序列和调节基因组成
结构基因:包括数个功能上有关联的基因,它们串联排列,共同构成编码区。这些结构基因共用一个启动子和一个转录终止信号序列,因此转录合成时仅产生一条mRNA长链,为几种不同的蛋白质编码。这样的mRNA分子携带了几个多肽链的编码信息,被称为多顺反子【polycistron】mRNA
调控序列:包括启动子【promote r】和操纵元件【operator】
启动子:RNA聚合酶和各种调控蛋白作用的部位,是决定基因表达效率的关键元件
操纵元件:是一段能被特异的阻遏蛋白识别和结合的DNA序列
调节基因【regulatory gene】 :编码能够与操纵序列结合的阻遏蛋白
阻遏蛋白的作用
识别、结合特异的操纵序列,抑制基因转录,所以阻遏蛋白介导负调节【negative regulation】。阻遏蛋白介导的负性调节机制在原核生物中普遍存在。
其它调控蛋白:特异因子,激活蛋白
特异因子决定RNA聚合酶对一个或一套启动序列的特异性识别和结合能力
激活蛋白可结合启动子邻近的DNA序列,提高RNA聚合酶与启动序列的结合能力,从而增强RNA聚合酶的转录活性,是一种正调控【positive regulation】。
分类
正转录调控
概念
在没有调节蛋白质(激活蛋白)存在时基因是关 闭的,加入这种调节蛋白质后基因活性就被开启
正控诱导:效应物分子(诱导物)的存在使激活蛋白处于活性状态
正控阻遏:效应物分子(辅阻遏物)的存在使激活蛋白处于非活性状态
负转录调控
在没有调节蛋白质(阻遏蛋白)存在时基因是表达的,加入这种调节蛋白质后基因表达活性便被关闭
负控诱导:阻遏蛋白与效应物(诱导物)结合时,结构基因转录
负控阻遏:阻遏蛋白与效应物(辅阻遏物)结合时结构基因不转录
乳糖操纵子
结构基因:Z、Y及A,分别编码β-半乳糖苷酶、透酶和乙酰基转移酶
调控区:操纵元件O、启动子P、分解代谢物基因激活蛋白(CAP,cAMP结合蛋白)结合位点
调节基因I:编码阻遏蛋白(与O序列结合,关闭操纵子)
乳糖操纵子受到阻遏蛋白和CAP的双重调节
阻遏蛋白的负性调节:无乳糖时,I序列表达的Lac阻遏蛋白与O序列结合,阻碍RNA聚合酶与P序列结合,抑制转录启动。
诱导物:别乳糖(由乳糖转变而来)
机理:别乳糖与阻遏蛋白结合,促使阻遏蛋白从O序列脱离,诱导基因表达
CAP的正性调节
正调节物:cAMP ,葡萄糖缺乏时,cAMP浓度升高
机理:cAMP与CAP结合形成复合物,促使CAP结合CAP位点,激活RNA聚合酶
协同调节
Lac阻遏蛋白负性调节与CAP正性调节两种机制协调合作
色氨酸操纵子
色氨酸操纵子通过阻遏作用抑制基因表达
细胞内无色氨酸时,阻遏蛋白不能与O序列结合,因此色氨酸操纵子处于开放状态,结构基因得以表达。
细胞内色氨酸的浓度较高时,色氨酸作为辅阻遏物与阻遏蛋白形成复合物并结合到O序列上,关闭色氨酸操纵子,停止表达用于合成色氨酸的各种酶。
生理意义:最大限度地减少能量消耗。
色氨酸操纵子通过衰减作用抑制基因表达
转录衰减【transcription attenuation】
使已经开始转录的mRNA合成终止的基因表达调节方式,称为转录衰减。这种作用是利用原核生物中转录与翻译过程偶联进行,翻译时先合成的一段前导序列L来实现的
前导序列L的结构特点
它可以转录生成一段长度为162bp、内含4个特殊短序列的前导mRNA;
其中序列1有独立的起始和终止密码子,可翻译成为一个有14个氨基酸残基的前导肽,它的第10位和第11位都是色氨酸残基;
序列1和序列2间、序列2和序列3间、序列3和序列4间存在一些互补序列,分别都可以形成发夹结构。形成发卡结构的能力依次是1/2发夹>2/3发夹>3/4发夹;
序列4的下游有一个连续的U序列,是一不依赖于ρ因子的转录终止信号。
转录衰减的机制
色氨酸的浓度较低时,前导肽的翻译因色氨酸量的不足而停滞在第10/11的色氨酸密码子部位,核糖体结合在序列1上,因此前导mRNA倾向于形成2/3发夹结构,转录继续进行
色氨酸的浓度较高时,前导肽的翻译顺利完成,核糖体可以前进到序列2,因此发夹结构在序列3和序列4形成,连同其下游的多聚U使得转录中途终止,表现出转录的衰减
转录衰减的生理意义
原核生物这种在色氨酸浓度高时,通过阻遏作用(粗调)和转录衰减机制(精调)共同关闭基因表达的方式,保证了营养物质和能量的合理利用。