导图社区 内质网
这是一篇关于内质网的思维导图,主要从离心分离技术得到颗粒型和光滑型内质网、研究历程、内膜系统、形态结构与类型等这些方面作了阐述,
细胞生物学中溶酶体知识点总结。主要从形态结构和化学组成、形成与成熟过程、类型、功能这几个方面作了阐述。
第二章:细胞的分子基础。主要从细胞的小分子物质、细胞大分子物质这两个大的部分做了详细的阐述,有不足的地方欢迎指正。
社区模板帮助中心,点此进入>>
论语孔子简单思维导图
《傅雷家书》思维导图
《童年》读书笔记
《茶馆》思维导图
《朝花夕拾》篇目思维导图
《昆虫记》思维导图
《安徒生童话》思维导图
《鲁滨逊漂流记》读书笔记
《这样读书就够了》读书笔记
妈妈必读:一张0-1岁孩子认知发展的精确时间表
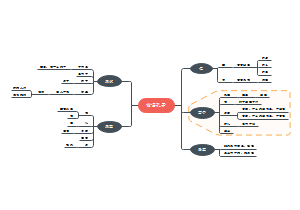
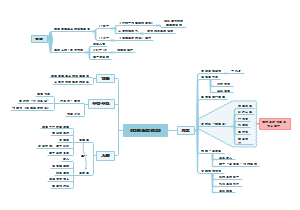
内质网
离心分离技术得到颗粒型和光滑型内质网(微粒体)
应用超速离心分级分离,从细胞浆中分离出直径100nm球囊状小泡,称生化分析等证明,微粒体由内质网和核糖体组成,具有内质网的基本功能。
研究历程
消失,进食后或活跃分泌时增多一一动质
1945年,电镜下--小泡小管、网状结构,命名内质网(ER)
内膜系统
细胞质中,结构、功能及其发生上相互密切关联的膜性结构细胞器之总称
包括
高尔基体
溶酶体
核膜
各种转运小泡
口诀:内高溶核转运泡
形态结构与类型
结构
分布
所有真核细胞胞质(成熟红细胞除外
基本结构单位
5-6nm的小管、小泡、扁囊
这些结构相互连通,构成连续的膜性三维管网结构系统。
位置
内质网向内与核膜沟通,向外与高尔基体、溶酶体等组分转换
形态结构、数量分布和发达程度--不同细胞或细胞的不同阶段各异,但不同物种的同类组织基本类似
类型
粗面内质网(RER
特点
多呈扁平囊状,参与外输性蛋白质及多种膜蛋白的合成、加工和转运
分泌肽类激素和蛋白的细胞中,RER高度发达;肿瘤细胞、未分化细胞则很少
肽激蛋白粗发达,肿瘤未分则很少。
特征
表面有核糖体附着
光面内质网(SER
与粗面内质网相通,是▲多功能细胞器;
★在不同细胞或不同生理时期,结构形态、分布及发达程度差别很大
呈表面光滑的管泡样网状形态结构
有的细胞以RER为主,有的以SER为主,随着生理状态改变,两者可以互相转换
衍生结构
实例
视网膜色素上皮细胞的髓样体
生殖细胞、快速增殖细胞、神经元与松果体细胞中的孔环状片层
神经松果生快殖,孔环片层细胞中。
来源
内质网局部分化、衍生而来的
化学组成
基本含量特征
常占细胞全部膜结构组成的50%,占细胞体积10%以上,相当于细胞质量的15%-20%
主要化学成分
膜脂:30%-40% 蛋白质:60%-70%
膜脂类
种类
磷脂(主要) 中性脂 缩醛脂 神经节苷脂等
磷脂比例**
卵磷脂55% 磷脂酰乙醇胺20~25% 磷脂酰肌醇和磷脂酰丝氨酸各占5~10% 鞘磷脂4~7%(大鼠肝细胞)
诸多酶系
葡萄糖-6-磷酸酶为主要标志性酶
分类
①与解毒功能相关的氧化反应电子传递酶系:细胞色素P450等
②与脂类物质代谢功能反应相关的酶类:脂肪酸CoA连接酶等
③与碳水化合物代谢功能反应相关的酶类:葡萄糖-6-磷酸酶
网质蛋白
内质网网腔中普遍存在的一类蛋白质(1、2、3、4、5皆属于分子伴侣)
共同特点
多肽链羧基端含有一个简称为KDLE或HDEL的4氨基酸序列驻留信号。 该信号可通过与内质网膜上相应受体的识别结合而驻留在内质网不被转运。
(1)免疫球蛋白重链结合蛋白:如BiP蛋白,阻止蛋白聚集或不可逆变性,协助蛋白折叠,与热激蛋白70(Hsp70)同源
(2)内质蛋白:内质网标志分子伴侣,参与新生肽链的折叠转运(又称葡萄糖调节蛋白94 GRP94)
(3)钙网蛋白:含钙结合位点,钙平衡调节、蛋白折叠加工、抗原递呈等方面发挥作用
(4)钙连蛋白:钙离子依赖的凝集素样伴侣蛋白,防止蛋白凝集和泛素化,阻止折叠不完全的蛋白离开内质网
(5)蛋白质二硫键异构酶(PDI):催化蛋白质二硫键交换,保证蛋白正常折叠
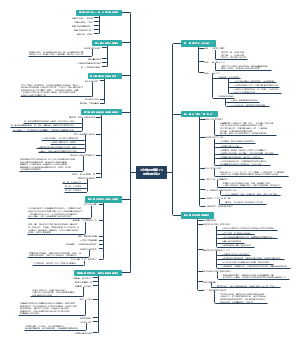
功能
糙面内质网主要功能
核糖体附着的骨架
作用
外输性蛋白质的合成、加工修饰、分选及转运
★参与蛋白质的合成
①分泌性蛋白:肽类激素、细胞因子、抗体、消化酶、细胞外基质蛋白等
②膜整合蛋白质:膜抗原、膜受体等
③可溶性驻留蛋白:定位于内质网、高尔基体、溶酶体等的可溶性驻留蛋白
信号肽指导的分泌蛋白质合成
★信号肽★
四种成分
信号肽
附着在内质网上的核糖体合成的肽链的N端的一段特殊氨基酸序列。一一疏水氨基酸序列,分泌蛋白肽链中普遍存在
细胞基质中信号识别颗粒SRP
向导作用
内质网膜上SRP受体
易位蛋白转运体(亲水蛋白通道)
信号肽假说 SPR是主角
step1
SRP由6个多肽亚单位和1个7S的小分子RNA构成复合体,SRP识别、结合新生信号肽,同时一端结合核糖体,形成SRP-核糖体复合结构,可使翻译暂停。
SPR识别&结合信号肽 另一端集合核糖体 →使翻译暂停
step2
与信号肽结合的SRP,识别、结合内质网膜上的SRP受体,并介导核糖体附着在内质网膜的转运体易位蛋白上。
SPR识别结合受体→ 介导核糖体附着在转易白
转运体外径8.5nm,中央孔2nm,SRP脱落(结合另外的新生肽链,重复利用),翻译重新启动。
SPR脱落→翻译继续
step3
在信号肽引导下,合成的肽链通过大亚基的中央管和转运体易位蛋白共同形成的通道进入内质网网腔。
肽链穿过通道进入网腔
信号肽切除,多肽转移完成后,转运体关闭。
信号肽切除→转移完成→转运体关闭
另外
核糖体结合是转运体开放的条件
转运体还介导损伤蛋白输出胞质
SPR-R也称停靠/船坞蛋白质
多肽核酸S P R,信号肽与核糖体,识别结合内受体,介导核体附转运,S P R 落翻译启,肽链通道进入腔。
新生分泌性蛋白质多肽链在细胞质基质中的游离核糖体上起始合成
新生多肽链的折叠与装配
★内质网腔中★
氧化型谷胱甘肽(GSSG) 协助二硫键的形成;
蛋白二硫键异构酶(网质蛋白) 促进二硫键的形成及多肽链的折叠
免疫球蛋白重链结合蛋(BiP)、内质蛋白、钙网蛋白及钙连蛋白等(网质蛋白/分子伴侣) 与折叠错误的多肽和未完成装配的蛋白亚单位识别结合;促进重新折叠、装配与运输
BiP属于热激蛋白70(Hsp70)家族成员,存在于真核细胞内质网,是一种“分子伴侣”---协助肽链的折叠组装和转运,但其本身不参与最终产物的形成。
热激蛋白也称作热休克蛋白
分子伴侣的共同特点:羧基基端有Lys-Asp-Glu-Leu(KDEL)--四氨基酸滞留信号肽,可结合于内质网膜受体蛋白,从而驻留于内质网腔,是一类驻留蛋白。
K D E L分子伴,四氨滞留信号肽,结合网膜受体P,驻留蛋白留网腔。
分子伴侣--质量监控因子
内质网腔中未折叠蛋白的积聚会促使内质网分子伴侣蛋白表达增加。
蛋白质的糖基化
糖基化:单糖或寡糖与蛋白质间,通过共价键结合成糖蛋白的过程
★粗面内质网中的糖基化:寡糖与蛋白质天冬酰胺残基侧链的氨基基团结合,即N-糖基化
糖基化需要一个共同的前体--多种单糖组成的14寡糖
催化糖基化的酶一一糖基转移酶,RER内膜整合蛋白
活化:寡糖与多贴醇分子连接
蛋白质的胞内运输
正确修饰加工的蛋白质,被内质网膜包裹,以“出芽”方式形成膜性小泡而转运
膜泡转运的两种形式
①形成转运小泡进入高体尔基复合体
②分泌蛋白以膜泡形式直接进入大浓缩泡,发育成酶原颗粒,排出细胞(仅存于某些哺乳类胰腺外分泌细胞)
反映蛋白质分泌特点:所有分泌蛋白质的胞内运输过程,始终以膜泡形式完全隔离于细胞质基质进行转运的
穿膜驻留蛋白插入转移
蛋白质分选的起始
信号斑是新生肽链合成后折叠形成的特定三维结构,其功能类似于信号肽,但通常会被保留
光面内质网主要功能
脂质的合成与转运
合成过程&催化酶
除线粒体特有的两种磷脂,几乎全部膜脂由内质网合成 一一底物来源于细胞质中,催化酶定位于内质网膜上
小肠吸收的甘油、甘油一酯、脂肪酸,在内质网中被重新合成甘油三酯
①脂酰基转移酶催化2分子脂酰辅酶A与甘油-3-磷酸反应,形成磷脂酸;
②磷酸酶催化磷脂酸脱磷酸,生成双酰甘油;
③胆碱磷酸转移酶催化双酰甘油添加极性基团,形成磷脂分子
合成的脂类常与糙面内质网合成的蛋白质结合成脂蛋白,经由高尔基体分泌出去;分泌出去后常运输血液中的胆固醇、甘油三酯等到脂肪组织
内质网向其它膜结构转运脂类的两种形式:
出芽形成小泡
①出芽小泡转运到高尔基体、溶酶体、质膜
磷脂交换蛋白
②以水溶性的磷脂交换蛋白为载体,与之结合形成复合体进入细胞质基质通过自由扩散,到达缺少磷脂的线粒体和过氧化物酶体膜上
糖原的代谢(分解过程)
肝细胞中光面内质网膜的葡萄糖-6-磷酸酶,催化糖原在胞质中降解的产物——葡萄糖-6-磷酸的去磷酸化;
去磷酸化的葡萄糖经由内质网,进入血液。
细胞解毒主要场所
地位
肝脏是外源性、内源性毒物及药物分解解毒的主要器官,其解毒作用主要由光面内质网来完成
原理
肝细胞光面内质网上,有丰富氧化和电子传递酶系:细胞色素P450;NADPH-细胞色素系列还原酶等
解毒机制:在电子传递的氧化还原过程中,催化多种化合物氧化或羟化,使毒物/药物被破坏;或增加了毒物/药物的极性,使之易于排泄
肌细胞钙离子的储存场所
肌细胞中发达的光面内质网特化为肌质网
肌浆网上Ca2+-ATP酶把胞质中的Ca2+泵入网腔储存;受细胞外信号作用,Ca2+向胞质中释放
钙离子浓度:胞质小于肌浆网/细胞外
肌浆网中存在大量钙结合蛋白,每个这样的蛋白可结合30个Ca2+
胃酸胆汁合成分泌
口诀:解毒代糖合脂质,胃酸胆汁钙离子。
内质网应激
▲