导图社区 蛋白质的生物合成
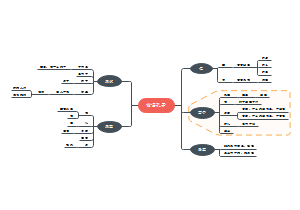
蛋白质的生物合成
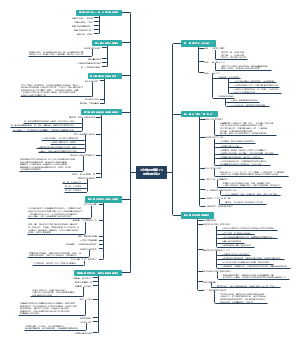
蛋白质是生命的物质基础,是构成细胞的基本有机物,是生命活动的主要承担者。本图梳理了蛋白质合成体系、氨基酸与tRNA的链接、肽链的合成过程、蛋白质合成的干扰与抑制,收藏下图学习吧!
编辑于2021-07-22 11:09:22- 蛋白质合成
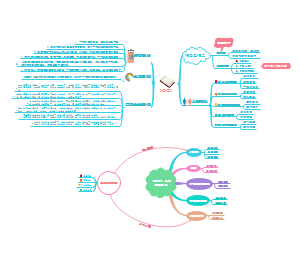
- 影像 中枢神经系统
《医学影像学》中枢神经系统思维导图,帮助大家更好滴掌握知识点,中枢神经系统(英文名称:Central Nervous System,中文名称中枢神经)是由脑和脊髓组成(脑和脊髓是各种反射弧的中枢部分),是人体神经系统的最主体部分。中枢神经系统接受全身各处的传入信息,经它整合加工后成为协调的运动性传出,或者储存在中枢神经系统内成为学习、记忆的神经基础。人类的思维活动也是中枢神经系统的功能。
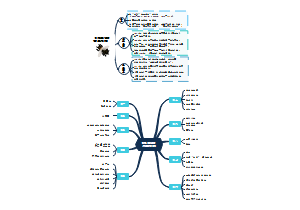
- 病原性真菌思维导图
医学微生物学里主要病原性真菌的知识点包括浅部感染真菌、深部感染真菌、皮下组织感染真菌、地方流行真菌,结构型知识框架方便学习记忆!
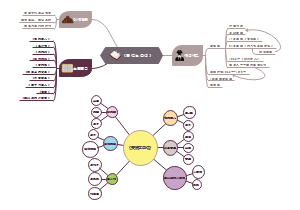
- 真菌学知识导图
真菌学是研究真菌形态、分类、生理、生态、发育和遗传的科学。它的基本任务是揭示真菌生存和发展的规律,进而获得利用、控制和改造真菌的知识和技能,有需要的收藏下图学习吧!
蛋白质的生物合成
社区模板帮助中心,点此进入>>
- 影像 中枢神经系统
《医学影像学》中枢神经系统思维导图,帮助大家更好滴掌握知识点,中枢神经系统(英文名称:Central Nervous System,中文名称中枢神经)是由脑和脊髓组成(脑和脊髓是各种反射弧的中枢部分),是人体神经系统的最主体部分。中枢神经系统接受全身各处的传入信息,经它整合加工后成为协调的运动性传出,或者储存在中枢神经系统内成为学习、记忆的神经基础。人类的思维活动也是中枢神经系统的功能。
- 病原性真菌思维导图
医学微生物学里主要病原性真菌的知识点包括浅部感染真菌、深部感染真菌、皮下组织感染真菌、地方流行真菌,结构型知识框架方便学习记忆!
- 真菌学知识导图
真菌学是研究真菌形态、分类、生理、生态、发育和遗传的科学。它的基本任务是揭示真菌生存和发展的规律,进而获得利用、控制和改造真菌的知识和技能,有需要的收藏下图学习吧!
- 相似推荐
- 大纲