导图社区 文献阅读
- 109
- 1
- 0
- 举报
文献阅读
文献阅读思维导图:包含研究内容,研究了这三种变化对烟草种子萌发的影响,天然烟草种子库的分离纯化,化学分析在康奈尔营养分析实验室使用标准程序进行。等等
编辑于2022-04-14 09:25:51- 文献阅读
- 相似推荐
- 大纲



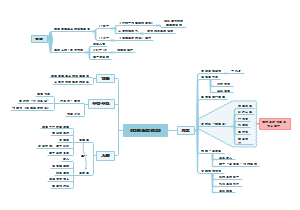

文献阅读
UP IN SMOKE: II. GERMINATION OF Nicotiana attenuata IN RESPONSE TO SMOKE-DERIVED CUES AND NUTRIENTS IN BURNED AND UNBURNED SOILS
研究内容
研究了这三种变化对烟草种子萌发的影响
火灾后的一年生植物被认为使种子库萌发与火灾后的环境同步,以响应:(1)与火灾有关的线索或(2)由火灾后营养物质矿化引起的营养物质供应速率的增加,(3)去除在燃烧前占据土壤的优势植被或与优势植被相关的微生物群落产生的化感物质 野生二倍体烟草种子库是由1993年采自4种优势灌木(Coleogyne ramosissima)、丝兰(Yucca baecata)、枸杞(Lycium andersonnii)和野生二倍体烟草种子库的焚烧和未焚烧土壤制成的。该地区在1992年被烧毁,并且至少自1988年以来,在两次干洗中,N. attenuata种群一直存在。
实验方法
土壤收集
从烧毁区内随机选取的4种优势灌木中的每一种植物下方取出大约50公斤的A ~层。从烧过的树桩或从黑化的树桩基部发芽的树叶中鉴定出物种。
A ~土层在本区灌丛之间几乎不存在,位于灌丛下直达5 ~ 10 cm厚
土壤样品也分别采自4种植物中的5种植物,分别采自邻近未燃区在燃烧区周长60 m以内但距离燃烧区不超过5 m的区域。
取上层5 ~ 10cm土层的土样,用塑料袋保存,用于化学分析和人工种子库试验
从发现大量植物(>75)生长的9个冲刷区收集了50公斤土壤
从带有小型和大型种子库(分别在图1中的 2和6)的冲洗土壤中取样进行化学分析,筛选以去除N. attenuata种子,并用于人工种子库实验。
天然烟草种子库的分离纯化
49个土壤采集物( 烧毁和未烧毁的场所和冲洗 )中的30公斤分成等份,摊成4个43厘米x20厘米的平地,平板布置成行排列。每个土壤采集2个托盘喷洒2升水,2个平板喷洒1:50稀释的液烟。每天浇水2次,连续20 d记录幼苗出苗数。
土壤养分分析
将1公斤的所有土壤分成一份,研磨成细粉(850目)。
化学分析在康奈尔营养分析实验室使用标准程序进行。分析样品的pH值(1:1 v/v土壤-0.01 M CaCI~悬浮液)、有机质(在500°C下燃烧2小时损失)和“有效”营养物质,用Morgan’s溶液(10%醋酸钠用1:5 v/v土壤溶液缓冲至pH值4.8)萃取烘干(110°C)样品。
采用原子吸收法测定K、Mg、Ca、Mn、Fe、A1和Zn;采用氯化亚锡和肼还原法分别比色法测定NO3中的P和N。
所有的营养价值都用毫克每公斤来表示。
人工种子库实验
用从田间收集的土壤创建了人工种子库,在这些土壤中添加了温室生长的具有高度休眠的植物的种子。
第一个实验
检测了燃烧后土壤中硝酸盐和发芽提示浓度增加的能力,以解释燃烧后土壤萌发潜力的增加。
采用3个处理组:(1)9.8 mM KNO3;(2) 1: 300 v/v液体烟9.8 mM KNO3;和(3)1:300 v/v液体烟在双蒸馏水
处理1和处理2的种子库双生率的对比描述了饱和硝酸盐浓度下添加线索的效果,处理2和处理3的对比描述了饱和硝酸盐浓度下添加线索的效果。 实验1包括42个土壤收集点(从烧过和未烧过的地方收集4个物种,外加2次洗涤)× 3个处理× 5个重复= 630个人工种子库。
第二个实验
增加饱和量的硝酸和提示未燃的土壤会增加这些土壤的发芽势,发现在同样的待遇燃烧土壤,然后开火的影响可以充分描述土壤的萌发潜力增加这些化合物。
采用2个处理组:(1)液体烟在完全营养液中稀释1∶300 v/v;(2)液体烟在双蒸馏水中稀释1∶300 v/v
完全营养液由9.8 mM KNO3在无氮营养液中组成。 无氮营养液由0.129 g/L CaSO4·2H20、0.123 g/L MgSO4·7H20、0.048 g/L K2HPO4、0.031 g/L KH2PO4、1.393 mg/L FeSO4·7H20、1.863 rag/L Na2EDTA·2H20和0.5 ml/L 微量营养液组成,由2.533 g/L H3BO3、1.634 g/L MnSO4·1H2O、0.151 g/L Na2MnO4·2H20、0.440 g/L ZnSO4·7H2O、 0.080 g/L CuSO4·5H20、 0.020 g/L CoCI 2· 6H20构成
在两个实验中,每种土壤类型的每种处理都准备了5个重复的种子库。每个复制的种子库包括10颗种子,放置在一个蛋奶酥杯(Solo 1 oz., P100)的6克干土壤表面。杯中加入3.0 ml适当的溶液。因此,在添加硝酸盐的处理中,除了土壤中已经存在的NO3外(NO3在0 ~ 0.2 mg之间),还添加了0.67 mg的NO3。杯子用透明盖子密封(Solo PL1盖子),放置在生长室(14L: 10D的光周期,200 u.M/m2/sec的PAR, 30°C日-22°C的夜间温度循环)
人工种子库每天在生长室内的不同位置轮流放置,在生长室内48小时后,每天检查种子的萌发情况,直到第7天。计算各种子库各计数下所有种子萌发率,并进行反正弦变换求正态。对转化百分比采用重复测量的三因素方差分析分析主效应,并对这些方差分析进行多因素对比,以检验特定的假设。采用X²试验,对烟熏处理土壤和对照土壤的自然衰减氮种子库的出苗数进行了分析。
发芽被定义为种子皮的分裂和胚根的出现,通过头戴的10倍放大镜观察。
研究结果
自然烟草种子库
与仅用水处理的土壤相比,烟熏处理的土壤中出现的幼苗数量显著增加(x² = 60.83p < 0.001)。
在洗涤中持续生长的植物产生的种子中,有一部分是休眠的,需要烟雾提示才能萌发。
土壤化学
通过双向方差分析,燃烧显著增加了所有物种采集的土壤中NO3 N、P、Mn、Ca和Mg的浓度(图2、表1)
燃烧增加了丝兰土壤中钾的浓度,但降低了枸杞土壤中钾的浓度,这是由显著的燃烧x种相互作用决定的(图1、表2)
收集的土壤之间的显著差异在不同的物种被发现P、K、钙、和镁浓度(平均±SEM)和土壤pH值(表1)
2次冲刷的土壤Mn、Zn、Cu和pH的浓度显著(Ps < 0.05)高于烧伤附近的土壤样品
人工种子库实验1
用烧过的土壤制成的人工种子库的种子发芽率始终高于用未烧过的土壤制成的种子库(图3,表2)
显著烧伤状态×物种和烧伤状态×物种×处理之间的交互作用也存在显著差异(表2)
通过三种重复测量方法(ANOVA)对这三个问题进行了多变量对比(表2)。在不同的物种和烧伤处理中,发现硝酸盐过量和硝酸盐过量两种线索的显著效应。证明这些土壤中不含有足以饱和发芽反应的大量提示和硝酸盐。
在未焚烧的丝兰土壤中添加饱和量的cue和硝酸盐能够提高其萌发率,但在统计学上差异不大(P=0.241;表2)从经过相似处理的烧丝兰土壤中得到。这一结果表明,焚烧对丝兰土壤的影响可以用土壤中硝酸盐含量的增加和提示来解释。但在黑灌丛、苦参灌丛和枸杞灌丛下,添加提示素和硝酸盐对土壤萌发率的影响不一致,仍显著降低(Ps < 0.003;表2)添加饱和量的cue和硝酸盐后,未燃烧的土壤比燃烧的土壤高。
人工种子库实验2
在洗涤中的萌发主要受土壤缺乏烟源萌发提示的限制
秸秆和全营养剂处理的火烧与未火烧苦灌丛土的发芽率差异不显著(P = 0.452;因此,火对苦灌丛下采集的土壤萌发势的影响可以用土壤养分的增加和提示来解释。然而,由于提示和营养物质的添加对黑灌丛和枸杞灌丛下的土壤没有相同的影响,我们认为这些变化不足以解释这两种植物被烧后土壤萌发势较高的原因。这一结果构成了这些灌木产生的化感因子的存在的间接证据,可以通过焚烧去除。
研究结论
与烟雾有关的抑制剂作的用必须小于未燃烧过的土壤中假定存在的抑制剂
燃烧过的土壤比未燃烧过的土壤萌发率更高
在特定的洗涤条件下,野生二倍体烟草(N. attenuata)种群的持久性可能是由于较低的草食率和较高的水分状况
与烟有关的萌发线索、营养和假定的化感化合物的破坏都可能参与种子库的火后萌发反应,但萌发线索可能是最重要的
原因:添加过量硝酸盐的萌发线索或完全营养物质比添加多因素对比确定的过量萌发线索的营养物质所占的方差更大。(实验1、2 )
只有萌发线索加入对从这些冲洗中取出的天然种子库产生了很大的响应,而用这些冲洗中的土壤制成的人工种子库对硝酸盐的加入没有明显的响应。
天然种子库:图1 人工种子库:图4
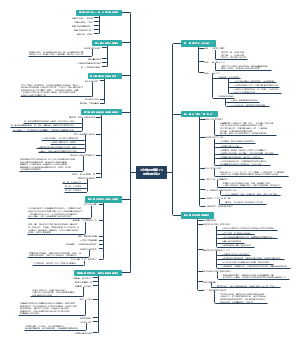
Evidence of different compounds in smoke derived from legumes and grasses acting on seed germination and seedling emergence
实验内容
使用生菜沙拉碗(Lactuca sativa)是一种快速的生物测定来追踪这三种烟熏溶液中的活性化合物。采用柱色谱法和高效液相色谱法对活性组分进行分离鉴定。
五种KAR1类似物(KAR2到KAR6),也被称为kararrikins,被发现并通过烟雾溶液的化学合成得到 植物源烟中的主要活性化合物是karikinolide(KAR1):3-methyl-2H-furo [2,3-c]-pyran-2-one KAR1可以刺激种子在非常低的浓度发芽,扩大环境条件下种子可以发芽和在增强杂草幼苗生长和药用植物发挥积极作用。
苜蓿(Medicago sativa, Fabaceae)产生的烟与草原干草(Festuca hallii,Poaceae)和小麦秸秆(Triticum aestivum,Poaceae)产生的烟对萌发的影响不同,说明不同的烟可能含有不同的活性化合物。
研究目的
(1)确定苜蓿、草原干草和小麦秸秆产生的烟中是否存在不同的活性化合物
(2)研究不同植物来源的烟和KAR1中的活性化合物如何与温度和光相互作用,影响羊茅普列里(Fescue Prairie)物种的种子萌发和幼苗生长
实验方法
烟和植物材料
18个不同品种的莴苣种子,清洁后的种子被保存在密封塑料袋中,并在萌发试验前存放在-20°C下。
发芽试验采用完全随机设计,共5个重复。30颗生菜的每个品种的种子被安置在10cm直径培养皿内衬两层Whatman 1号滤纸和5ml蒸馏水滋润下安全的绿灯,然后孵化25°C下12h 光照-12h 黑暗,或者24h黑暗。培养皿置于透明密封袋中,在光下孵育12h,在暗下孵育12h。用两层铝箔包裹的密封袋来容纳培养皿,种子在黑暗中孵育24h。孵育1天后记录萌发情况。
栽培品种(L. sativa L. cv. )以光、暗发芽差异最大的沙拉碗(Salad Bowl)为试验品种,对烟熏类型进行比较。
沙拉碗生菜生物测定
对于每个生物测定试验,采用完全随机设计,有5个重复。将30颗沙拉碗生菜种子放在10cm的培养皿中,培养皿内衬两层Whatman 1号滤纸,在黑暗的绿色安全灯下用5ml测试溶液润湿。每次实验均以蒸馏水为对照,各分离馏分采用不同的稀释度以保证活性的最佳浓度范围。培养皿密封在两层铝箔包裹的塑料袋中,在25°C的黑暗中孵育。孵育1天后记录萌发情况。
烟溶液的分馏
从苜蓿、草原干草和小麦秸秆产生的烟溶液中分离活性化合物的方法是基于Flematti等人(2008)的修改。
从紫花苜蓿、草原干草和麦秸中产生的每种库存烟雾溶液共2L,过滤(32cm,Whatman 1号滤纸)并分离。每升烟雾溶液用乙酸乙酯(3×200mL)彻底萃取。然后使用NaOH水溶液(1%w/v)(5×100 mL)将合并的有机提取物溶液分馏成酸(NaOH可溶)和中性组分。所得中性组分溶液用Na2SO4干燥,过滤,并在真空中蒸发以除去水分,并得到中性组分(452 mg苜蓿,422 mg草原干草,428 mg小麦)
浓缩的中性组分使用填充有50 g硅胶60(Merck,0.040–0.063 mm)的2.5×30 cm柱进行柱层析,并用己烷:乙酸乙酯梯度洗脱[己烷比例:100,90,80,70,60,50,40,30,20,10和0%(v/v);每种混合物150 ml等份]。在真空中蒸发活性组分,然后将其置于反相(RP)C18柱(Waters-Sep-Pak 12 cc Vac柱)中,用水:甲醇梯度洗脱[甲醇比例:0,10,15,20,25和100%(v/v);每种混合物50 ml等份]。活性组分在真空中蒸发至1.5 ml。
使用C18-RP高效液相色谱柱(Chromolith Performance RP-18e 100-4.6)分析苜蓿、草原干草或麦秸烟雾溶液中的部分活性组分(20µl),用乙腈:水梯度指定为7%–14%–95%–7%–7%乙腈/水,在0–14–15–16–20分钟内进行进一步分离。洗脱速率为2ml/min。在330nm处测量UV吸光度。根据洗脱模式,在3-12.5分钟(3-5.7、5.7-6.6、6.6-9、9-10、10-11和11-12.5分钟)之间收集组分。20µl纯KAR1(0.1 mg/ml)样品用与标准品相同的乙腈基方法洗脱。通过每次将(RP)C18柱层析后获得的80µl馏分应用于C18-RP HPLC,使用上述相同的方法进行10次分馏,可获得更多活性馏分。从苜蓿、草原干草和麦草产生的烟雾溶液中,总共获得了18毫升(2毫升/分钟,每次0.9分钟,每次10次)的活性组分。
羊茅普列里植物种类的选择
选择了四种来自克伦草原的菊植科物,其中总幼苗密度在燃烧后显著增加,包括流苏鼠尾草(Artemisia frigida)、白蒿(Artemisia ludoviciana)、加拿大蓟(Cirsium arvense)和加拿大马鞭草(Conyza canadensis)。在室温和环境相对湿度下成熟一个月后,清洁的种子保存在密封的塑料袋中,并在-20°C下储存,直到用于发芽和幼苗生长试验。
活性组分对种子萌发和幼苗生长的激发效应
将从苜蓿、草原干草或麦秸产生的烟雾溶液中获得的每个活性组分(18 ml)在1.07L 蒸馏水中稀释,相当于每2L 1/1 v/v储备烟雾溶液中活性化合物的浓度(2L×0.8÷1.5,因为对于每种类型的烟雾溶液,在反相色谱后总共获得1.5ml的部分,其中0.8ml经过HPLC进一步分离)。
为了确定KAR1对种子萌发的影响,将浓度为10﹣6 M of KAR1溶解在蒸馏水中,用作1/1 v/v KAR1溶液。四种不同的溶液(1/1 v/v),包括从紫花苜蓿、草原干草、麦秸和KAR1溶液中获得的活性组分,每种溶液都被制成三个系列稀释液,包括1/1000 v/v、1/100 v/v和1/10 v/v。用蒸馏水浸泡的种子作为对照。每一部分(18毫升)含有10%的乙腈。
为了消除乙腈对种子萌发的影响,将1.8ml乙腈溶解在1.07L蒸馏水中,视为1/1 v/v乙腈溶液。各种浓缩乙腈溶液(1/1000 v/v、1/100 v/v、1/10 v/v和1/1 v/v)对任何受试物种的萌发或幼苗生长没有影响。
每种植物的50颗种子被计数并放置在一个50ml的离心管中,垂直储存在穿孔纸盒中。种子在加入10毫升蒸馏水、1/1000 v/v、1/100 v/v、1/10 v/v或1/1 v/v每种水溶液或KAR1溶液后浸没。每个离心管用盖子密封,并在20°C的条件下黑暗24h。然后将种子转移到10cm长的培养皿中,培养皿内衬两层Whatman 1号滤纸,并在20°C的条件下黑暗干燥1周。然后用5ml蒸馏水湿润每个皮氏培养皿中的种子和滤纸,并在10/0°C或25/15°C条件下,在12h光照-12h黑暗或24h黑暗条件下培养。将皮氏培养皿放在透明的拉链袋中,供那些在光照12h-暗照12h的培养皿使用。用两层铝箔包裹的拉链袋用于放置在24h黑暗中孵化的种子的研究培养皿。
10/0°C或25/15°C条件下,在12h光照-12h黑暗或24h黑暗条件模拟了克伦草原春季(4月)和夏季(7月)的日平均温度(加拿大环境部)。
每周对冷地黄花、绿地黄花、阿尔文氏黄花和加拿大黄花的发芽情况进行统计,连续7周。有胚根的种子≥1mm被认为是发芽的。加入蒸馏水以保持滤纸湿润。在绿色安全光下检查24h黑暗处理中的种子发芽情况。每周对发芽的种子进行计数,并将其转移到一个新的培养皿中,让幼苗在相同温度下,在光照12h-黑暗12h的条件下生长7天,以进行种子发芽。7天后测量胚根和下胚轴的长度。
当测量时,幼苗可能在7到13天之间。
对每一个物种采用随机完全区组设计,对四种不同类型(苜蓿、草原干草、麦秸和KAR1)分别采用五种启动处理(1/1000 v/v、1/100 v/v、1/10 v/v、1/1 v/v和蒸馏水),并在四种发芽条件下各进行四次重复,其中包括10/0°C 12h光照-黑暗12h,10/0°C时黑暗24h,25/15°C光照12h–黑暗12h,25/15°C 黑暗24h。该实验重复一次。
数据分析
在使用t检验或方差分析进行分析之前,所有发芽数据均经过反正弦平方根转换。 不符合正态性假设的幼苗长度数据在分析之前进行了对数转换。 Shapiro–Wilk检验用于测试转换前后的数据正态性。转换后所有数据均为正态分布。治疗手段在P≤ 0.05.
用t检验比较25°C光照和黑暗条件下生菜品种间的发芽差异。采用单因素方差分析( ANOVA)研究了不同组分或烟熏类型对沙拉碗生菜种子分离后萌发的影响。在P≤0.05时采用Tukey’s检验分离治疗手段。
为了研究分离组分和KAR1对种子萌发和幼苗生长的影响,采用SAS 9.3版软件(SAS Institute Inc.,USA)中的混合模型程序,采用随机完全区组设计,分两次(每次重复四次)分析总发芽率、胚根长度、下胚轴和总根数。
对于每个处理中的每个物种、研究烟雾类型的启动效应被用作自变量。重复、区块和运行被视为随机效应。
研究结果
基于光响应的苜蓿品种筛选
在恒定25°C的条件下,在12小时光照-12小时黑暗和24小时黑暗中测试了18个不同品种的苜蓿的种子萌发(图1)。沙拉碗的发芽生菜在光照和暗照下的发芽率差异最大(P<0.01)。
不同烟雾溶液对沙拉碗莴苣种子萌发的影响
用草原干草和麦秆产生的1/5000 v/v烟雾稀释液处理后,沙拉碗莴苣的发芽率提高。与对照相比,不同稀释度的苜蓿烟对沙拉碗生菜的发芽没有影响(P=0.44)。
烟雾溶液对沙拉碗莴苣发芽的刺激作用因烟雾类型和稀释度而异(图2)
用沙拉碗生物测定法追踪植物烟雾中的活性化合物
草原干草和小麦秸秆乙酸乙酯萃取物启动后,沙拉碗生菜种子萌发率显著提高(P < 0.01)。苜蓿溶液乙酸乙酯部位对生菜种子萌发无影响。不同烟熏类型的3种水分组分对生菜种子萌发均无影响(P = 0.70)。
乙酸乙酯将苜蓿、草原干草或小麦秸秆制成的原液烟溶液分离成水(无机)和乙酸乙酯(有机)馏分(图3A)。 这两个馏分都用沙拉生菜生物测定法在一系列稀释下进行了测试,以确保活性的正确浓度范围。
与对照相比,草原干草和麦草溶液中的NaOH组分显著提高了种子发芽率(P<0.01)。然而,与对照相比,苜蓿中的NaOH组分对种子萌发没有影响。与对照相比,来自不同烟雾类型的三种酸性组分均未对沙拉碗生菜的种子萌发产生影响(P=0.10)。
使用NaOH将有机提取物分为酸性和中性组分(图3B)。这两种组分均通过沙拉碗生菜生物测定法在一系列稀释液中进行测试,以确保活性的正确浓度范围。
浓缩的中性部分通过色谱分离,用乙酸乙酯:己烷梯度洗脱(乙酸乙酯比例:0,10,20,30,40,50,60,70,80,90和100%)。用沙拉碗生菜生物测定法在一系列稀释液中测试不同的组分,以确保活性的正确浓度范围。在70:30的乙酸乙酯:己烷溶液中洗脱的具有发芽刺激活性的部分出现在所有三种烟雾类型中(图4)。
通过反相色谱分离70:30乙酸乙酯:己烷部分,用甲醇:水梯度(甲醇比例:0,10,15,20,25和100%)洗脱。然后用沙拉碗生菜生物测定法在一系列稀释液中测试不同的组分,以确保活性的正确浓度范围。具有发芽刺激活性的组分在10:90甲醇:水中洗脱,并从草原干草和麦秸中获得(图5)。与对照组相比,苜蓿中的任何一种成分都不影响沙拉碗生菜的种子萌发(P=0.31)。
使用HPLC分离10:90甲醇:水部分。根据洗脱模式,在3到12.5分钟(3-5.7、5.7-6.6、6.6-9、9-10、10-11和11-12.5分钟)之间收集六个部分,分别视为第1到第6部分。用沙拉碗生菜生物测定法在一系列稀释液中测试不同部分,以确保活性的正确浓度范围。用草原干草和麦秸的第二部分处理种子,可显著提高种子发芽率(P<0.01)(图6)。与对照相比,苜蓿中的六种成分均未对沙拉碗生菜的种子萌发产生影响(P=0.61)。
KAR1与从草原干草和麦秸产生的烟雾中获得的组分中的活性化合物共洗脱,但不与从苜蓿中获得的组分共洗脱(图7)。
活性组分和KAR1对所选物种种子萌发和幼苗生长的影响
A. frigida
在10/0°C条件下,在光照12h-黑暗12h的条件下,草原干草溶液中的活性组分(P<0.01)和KAR1(P<0.01)的启动效应影响冷蒿的总种子萌发(表1)。用草原干草中的1/1 v/v活性组分引发,提高种子发芽率,提高。用1/10 v/v和1/1 v/v KAR1溶液引发种子,使种子发芽率分别提高。与对照相比,不同稀释度的麦秸(P=0.09)和苜蓿(P=0.3)引发对冷青蒿的萌发没有影响。冷蒿种子胚根、下胚轴和幼苗总长度在任何研究的萌发条件下均不受任何研究溶液引发效应的影响。
A.ludoviciana
草原干草(P=0.04)、小麦秸秆(P=0.04)和KAR1溶液(P=0.02)中的活性组分在25/15°C的24h黑暗条件下的启动效应影响了A.ludoviciana的总种子萌发(表1)。用草原干草和麦秸中的1/1 v/v组分引发,种子发芽率提高。用1/1 v/v KAR1溶液引发后,种子发芽率提高。与对照相比,不同稀释度的苜蓿组分(P=0.25)中的引发剂对A.ludoviciana的萌发没有影响。在任何研究的发芽条件下,任何研究溶液的任何引发效应均不会影响A.ludoviciana的种子胚根、下胚轴和总幼苗长度。
C. canadensis
草原干草(P=0.02)、小麦秸秆(P=0.03)和KAR1溶液中的活性组分引发效应影响加拿大一枝黄花的总种子萌发(P=0.02)在25/15°C的条件下,在12h的光照-12h的黑暗中(表1)。用草原干草中的1/10 v/v组分引发,种子发芽率提高。用1/1000 v/v小麦组分引发,使种子发芽率提高。用1/1000 v/v和1/100 v/v的KAR1溶液引发后,种子发芽率提高。与对照相比,不同稀释度的紫花苜蓿(P=0.43)对引发对加拿大一枝黄花萌发无影响。在任何研究的发芽条件下,加拿大一枝黄花的种子胚根、下胚轴和幼苗总长度均不受任何研究溶液的任何引发效应的影响。
C. arvens
在25/15°C条件下,光照12h-暗照12h,KAR1溶液的引发效应影响了卷耳的胚根长度(表2)。用1/1 v/v KAR1溶液打底(P<0.01)显著减少胚根长度。与对照组相比,苜蓿(P=0.28)、草原干草(P=0.06)和麦秸(P=0.46)的不同稀释液中的引发剂对卷耳的胚根长度没有影响。 在任何研究的发芽条件下,任何研究溶液的任何引发效应均不会影响卷耳的总种子发芽率、下胚轴和总幼苗长度。
表1
表2
研究结论
KAR1并不普遍存在于烟雾溶液中
KAR1有利于羊茅普列里本地物种的种子萌发