导图社区 道德经读书笔记守道和修德
- 92
- 4
- 4
- 举报
道德经读书笔记守道和修德
道德经读书笔记守道和修德,修德方能明道,明道才能守道,希望这份脑图会对你有所帮助。
编辑于2023-05-22 14:40:19 广东- 相似推荐
- 大纲
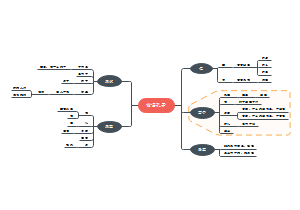



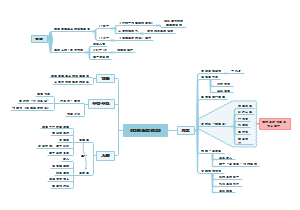
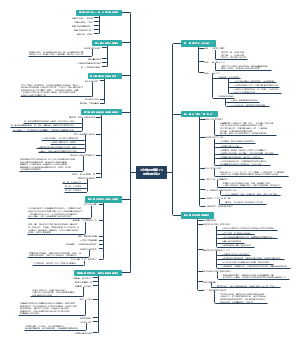

道德经
守道
守道
故常无欲,以观其妙,常有欲,以观其徼。
故常无欲,以观其妙,
因此,要常从“无”中去观察领悟“道”的奥妙
常有欲,以观其徼。
要常从“有”中去观察体会“道”的端倪。
端倪,汉语词语,拼音是duān ní,意思是事情的眉目, [1] 出自《庄子·大宗师》。
是以圣人处无为之事,行不言之教,万物作焉而不辞,生而不有,为而不恃,功成而弗居。
因此圣人用无为的观点对待世事,用不言的方式施行教化:
听任万物自然兴起而不为其创始,
创始,指创建;创立。有时指在原来一种物质或方法基础上,通过丰富的想象和发明创造出一种新兴的文化或科技即为创始。语出晋 陆机 《羽扇赋》:“夫创始者恒朴,而饰终者必妍。”
有所施为,但不加自己的倾向,功成业就而不自居。
行不言之教
信不足,焉有不信。
统治者的诚信不足,就会有人不信任。
故同于道者,道亦得之;同于失,道亦失之。
。同于道的人,道也乐于得到他;同于德的人,德也乐于得到他;同于失的人,失也乐于得到他。
故从事于道者同于道,德者同于德,失者同于失。
所以,从事于道的就同于道,从事于德的就同于德,从事于失的人就同于失。
故飘风不终朝,骤雨不终日。
狂风刮不了一个早晨,暴雨下不了一整天。
天地,天地尚不能久,而况于人乎?
天地的狂暴尚且不能长久,更何况是人呢?
希言自然。
不言政令不扰民是合乎于自然的。
不尚贤,使民不争;不贵难得之货,使民不为盗;不见可欲,使民心不乱。
不尚贤,使民不争;
不推崇有才德的人,导使老百姓不互相争夺;
不贵难得之货,使民不为盗;
不珍爱难得的财物,导使老百姓不去偷窃;
不见可欲,使民心不乱。
不显耀足以引起贪心的事物,导使民心不被迷乱。
是以圣人之治,虚其心,实其腹,弱其志,强其骨,常使民无知无欲。
因此,圣人的治理原则是:排空百姓的心机,填饱百姓的肚腹,减弱百姓的竞争意图,增强百姓的筋骨体魄,经常使老百姓没有智巧,没有欲望。
虚其心,
排空百姓的心机
实其腹
填饱百姓的肚腹
弱其志
减弱百姓的竞争意图,?这个翻译是否合适
志
“志”通常被认为是形声字。金文(图1、2)和小篆(图3)的“志”字,下部的“
”字是形符,表示与人的心理和思想活动有关。上面的字形是“
”字,作声符。其实“之”字也有表意作用。“之”来源于表运动、活动之意的“止”。指明内心的趋向为志。若据此,则“志”为形声兼会意字。早期文字到小篆,均为上“之”、下“心”;隶楷文字渐变“之”为“士”。
“志”本义为意念、心情。当某种意念、心情强化时,就成为愿望、志向,这是“志”意义的引申,也是“志”最常用的意义。“志”由“志向”又引申出许多意义。第一,由名词转为动词,即“向慕”“有志于”“期望”;第二,指神志。第三,指德行,因为德行与志向是密切相关的。第四,人有志向,就有了奋斗目标,好比射箭有了靶子,因此,“志”又指“目标”。第五,“志”是一个人思想精神的集中表现,因此就成为表明某一特征的记号,即标记、标志。“标记”又引申为“记”“记忆”,如博闻强志,即见闻广博,记忆力强。又引申为记载,记录,“志怪”即记述怪异之事,后在古典小说中形成了一个类别。“志”又可作名词,指记事的文章、著作,如“日志”“杂志”“墓志”“县志”。与“标志”的意义有关,“志”还与“帜”“痣”二字相通。旗帜是一个集团的标志,故“志”与“帜”相通。
在古代汉语中,上述标志和表示记忆、记录的“志”常与“识”(zhì)相通。《周礼·春官·保章氏》:“掌天星,以志星辰日月之变动。”注:“志,古文识。识,记也。”其实“志”与“识”原本是不同的词。
认为,“识”的本义就是“记”;而“志”的标记、记录的意义,如前所述,是由志向、心志引申出来的。所以“志”“识”是两词两字,而非两词一字。另一方面,“志”“识”也不是假借的关系,两字在上古的读只是接近,而非相同。但因两字音近义近,故可相通。
上述标志、记忆、记录之“志”后来又产生了一个分化字“誌”(图9),“誌”指标志、记忆、记录、记事的文章、书籍。因“誌”字的功能包括在“志”字之中,所以《第一批异体字整理表》将“誌”作为“志”的异体字处理,不再使用。
强其骨,
增强百姓的筋骨体魄,
常使民无知无欲
经常使老百姓没有智巧,没有欲望。
使夫知者不敢为也。
致使那些有才智的人也不敢妄为造事。
天地不仁,以万物为刍狗,圣人不仁,以百姓为刍狗。
天地不仁,以万物为刍狗,
天地是无所谓仁慈的,它没有仁爱,对待万事万物就像对待刍狗一样,任凭万物自生自灭
圣人不仁,以百姓为刍狗。
圣人也是没有仁受的,也同样像刍狗那样对待百姓,任凭人们自作自息。
刍狗
古时结草为狗形,以供祭祀之用,用毕则弃之。后借以比喻无用之物。
刍狗是一个汉语词语,典故名,典出
①刍(chu)狗:用草扎成的狗。古代专用于祭祀之中,祭祀完毕,就把它扔掉或烧掉。比喻轻贱无用的东西。在本文中比喻:天地对万物,圣人对百姓都因不经意、不留心而任其自长自消,自生自灭。正如元代吴澄据说:“刍狗,缚草为狗之形,祷雨所用也。既祷则弃之,无复有顾惜之意。天地无心于爱物,而任其自生自成;圣人无心于爱民,而任其自作自息,故以刍狗为喻。”
橐籥(tuó yuè)
虚而不屈,动而愈出。
它空虚而不枯竭,越鼓动风就越多,生生不息
多言数穷,不如守中。
政令繁多反而更加使人困惑,更行不通,不如保持虚静。
居善地,心善渊,与善仁,言善信,正善治,事善能,动善时。
居善地,
最善的人,居处最善于选择地方,
心善渊,
心胸善于保持沉静而深不可测,
与善仁,
待人善于真诚、友爱和无私,
言善信
说话善于恪守信用,
正善治,
为政善于精简处理,能把国家治理好
事善能
处事能够善于发挥所长
动善时
行动善于把握时机
载营魄抱一,能无离乎?
精神和形体合一,能不分离吗?
专气致柔,能如婴儿乎?
聚结精气以致柔和温顺,能像婴儿的无欲状态吗?
涤除玄览,能无疵乎?
洗清杂念而深入观照,能没有瑕疵吗?
爱民治国,能无为乎?
"爱民治国能遵行自然无为的规律吗?
天门开阖,能为雌乎?
感官与外界的对立变化相接触,能宁静吧?
明白四达,能无知乎?
明白四达,能不用心机吗?
生之畜之,生而不有,为而不恃,长而不宰,是谓玄德。
让万事万物生长繁殖,产生万物、养育万物而不占为己有,作万物之长而不主宰他们,这就叫做“玄德”。
玄德
玄德,读音为xuán dé,是一个汉语词语,意思是潜蓄而不著于外的德性。
埏埴以为器,当其无,有器之用。
揉和陶土做成器皿,有了器具中空的地方,才有器皿的作用。
凿户牖以为室,当其无,有室之用。
开凿门窗建造房屋,有了门窗四壁内的空虚部分,才有房屋的作用。
故有之以为利,无之以为用。
所以,“有”给人便利,“无”发挥了它的作用。
五色令人目盲,五音令人耳聋,五味令人口爽,驰骋畋猎令人心发狂,难得之货令人行妨。
五色令人目盲,
缤纷的色彩,使人眼花缭乱;
五音令人耳聋,
嘈杂的音调,使人听觉失灵;
五味令人口爽
丰盛的食物,使人舌不知味;
驰骋畋猎令人心发狂
纵情狩猎,使人心情放荡发狂;
难得之货令人行妨。
稀有的物品,使人行为不轨。
是以圣人为腹不为目,故去彼取此。
因此,圣人但求吃饱肚子而不追逐声色之娱,所以摒弃物欲的诱惑而保持安定知足的生活方式。
宠辱若惊,贵大患若身。
受到宠爱和受到侮辱都好像受到惊恐,把荣辱这样的大患看得与自身生命一样珍贵
宠为下。
得宠是卑下的
得之若惊,失之若惊,是谓宠辱若惊。
得到宠爱感到格外惊喜,失去宠爱则令人惊慌不安。这就叫做得宠和受辱都感到惊恐。
故贵以身为天下,若可寄于天下,爱以身为天下者,若可托天下。
所以,珍贵自己的身体是为了治理天下,天下就可以托付他;爱惜自己的身体是为了治理天下,天下就可以依靠他了。
古之善为士者,微妙玄通,深不可识。
古时候善于行道的人,微妙通达,深刻玄远,不是一般人可以理解的
夫唯不可识,故强为之容:
正因为不能认识他,所以只能勉强地形容他说:
豫兮,若冬涉川;犹兮,若畏四邻;俨兮,其若客;涣兮,若冰之将释;敦兮,其若朴;旷兮,其若谷;浑兮,其若浊。
豫兮,若冬涉川;
他小心谨慎啊,就像冬天里涉水过河;
犹兮,若畏四邻;
他警觉戒备啊,好像防备着邻国的进攻
俨兮,其若客;
他恭敬郑重啊,好像要去赴宴做客;
涣兮,若冰之将释;
他行动洒脱啊,好像冰块缓缓消融;
敦兮,其若朴;
他纯朴厚道啊,好像没有经过加工的原料;
旷兮,其若谷;
他旷远豁达啊,好像深幽的山谷;
浑兮,其若浊。
他浑厚宽容,好像不清的浊水。
孰能浊以止,静之徐清?
谁能使浑浊安静下来,慢慢澄清?
孰能安以久,动之徐生?
谁能使安静变动起来,慢慢显出生机?
保此道者不欲盈,夫唯不盈,故能敝而新成。
保持这个“道”的人不会自满。正因为他从不自满,所以能够去故更新。
致虚极,守静笃,万物并作,吾以观其复。
致虚极,守静笃,
尽力使心灵的虚寂达到极点,使生活清静坚守不变。
万物并作,吾以观其复。
万物都一齐蓬勃生长,我从而考察其往复的道理。
夫物芸芸,各复归其根。
那万物纷纷芸芸,各自返回它的本根。
复命曰常,知常曰明,不知常,妄作,凶。
复归于生命就叫自然,认识了自然规律就叫做聪明,不认识自然规律的轻妄举止,往往会出乱子和灾凶。
归根曰静,是谓复命。
返回到它的本根就叫做清静,清静就叫做复归于生命。
知常容,容乃公,公乃全,全乃天,天乃道,道乃久,没身不殆。
认识自然规律的人是无所不包的,无所不包就会坦然公正,公正就能周全,周全才能符合自然的“道”,符合自然的道才能长久,终身不会遭到危险。
知常容,
容乃公
公乃全
全乃天
天乃道
道乃久
没身不殆。
终身不会遭到危险。
符合自然的道才能长久,
周全才能符合自然的“道”,
周全才能符合自然的“道”,
公正就能周全,
无所不包就会坦然公正,
认识自然规律的人是无所不包的,
绝圣弃智,民利百倍,绝仁弃义,民复孝慈,绝巧弃利,盗贼无有。
绝圣弃智,民利百倍,
抛弃聪明智巧,人民可以得到百倍的好处;
绝仁弃义,民复孝慈,
抛弃仁义,人民可以恢复孝慈的天性;
绝巧弃利,盗贼无有。
抛弃巧诈和货利,盗贼也就没有了。
此三者以为文不足,故令有所属,见素抱朴,少私寡欲。
圣智、仁义、巧利这三者全是巧饰,作为治理社会病态的法则是不够的,所以要使人们的思想认识有所归属,保持纯洁朴实的本性,减少私欲杂念,
绝学无忧。
抛弃圣智礼法的浮文,才能免于忧患
曲则全,枉则直;洼则盈,弊则新;少则得,多则惑,是以圣人抱一为天下式。
曲则全,枉则直;
委曲便会保全,屈枉便会直伸;
屈枉,汉语词语,读音qū wang,意思是冤枉。
洼则盈,弊则新;
低洼便会充盈,陈旧便会更新;
少则得,多则惑,是以圣人抱一为天下式。
少取便会获得,贪多便会迷惑。所以有道的人坚守这一原则作为天下事理的范式,
不自见,故明;不自是,故彰;不自伐,故有功;不自矜,故长。
不自见,故明;
不自我表扬,反能显明;
不自是,故彰;
不自以为是,反能是非彰明;
不自伐,故有功;
不自己夸耀,反能得有功劳;
不自矜,故长。
不自我矜持,所以才能长久。
矜持是一个汉语词语,读音为jīn chí,意思是(1)局促;拘束(2)自鸣得意;自负(3)恪守;坚持正道。
企者不立,跨者不行,自见者不明,自是者不彰。
物或恶之,故有道者不处也。
因为它们是令人厌恶的东西,所以有道的人决不这样做。
企者不立,
踮起脚跟想要站得高,反而站立不住;
跨者不行,
迈起大步想要前进得快,反而不能远行。
自见者不明
自逞已见的反而得不到彰明;
自是者不彰。
自以为是的反而得不到显昭;
自伐者无功,自矜者不长。
自我夸耀的建立不起功勋;自高自大的不能做众人之长。
轻则失本,躁则失君。
轻率就会失去根本;急躁就会丧失主导。
善行无辙迹;善言无瑕谪;善数不用筹策;善闭,无关楗而不可开;善结,无绳约而不可解。
善行无辙迹;
善于行走的,不会留下辙迹;
善言无瑕谪;
善于言谈的,不会发生病疵;
疵病是汉语词汇,拼音cī bìng,释义为缺点,毛病;非议,指出缺点、毛病。出自《书·大诰》。
善数不用筹策;
善于计数的,用不着竹码子;
善闭,无关楗而不可开
善于关闭的,不用栓梢而使人不能打开;
善结,无绳约而不可解。
善于捆缚的,不用绳索而使人不能解开。
是以圣人常善救人,故无弃人;常善救物,故无弃物,是谓袭明。
是以圣人常善救人,故无弃人;
因此,圣人经常挽救人,所以没有被遗弃的人;
常善救物,故无弃物,是谓袭明。
经常善于物尽其用,所以没有被废弃的物品。这就叫做内藏着的聪明智慧。
故善人者,不善人之师;不善人者,善人之资。
故善人者,不善人之师;
所以善人可以做为恶人们的老师,
不善人者,善人之资。
不善人可以作为善人的借鉴。
不贵其师,不爱其资,虽智大迷。
不尊重自己的老师,不爱惜他的借鉴作用,虽然自以为聪明,其实是大大的糊涂。
知其雄,守其雌,为天下谿。
谿,汉语三级字, [1] 读作谿(xī ),同“溪”。溪字意指,山里的小河沟,泛指小河沟。
深知什么是雄强,却安守雌柔的地位,甘愿做天下的溪涧。
为天下谿,常德不离,复归于婴儿。
甘愿作天下的溪涧,永恒的德性就不会离失,回复到婴儿般单纯的状态。
知其白,守其黑,为天下式。
深知什么是明亮,却安于暗昧的地位,甘愿做天下的模式。
为天下式,常德不忒,复归于无极。
甘愿做天下的模式,永恒的德行不相差失,恢复到不可穷极的真理。
知其荣,守其辱,为天下谷。
深知什么是荣耀,却安守卑辱的地位,甘愿做天下的川谷。
为天下谷,常德乃足,复归于朴。
甘愿做天下的川谷,永恒的德性才得以充足,回复到自然本初的素朴纯真状态。
朴散则为器,圣人用之,则为官长,故大制不割。
朴素本初的东西经制作而成器物,有道的人沿用真朴,则为百官之长,所以完善的政治是不可分割的。
将欲取天下而为之,吾见其不得已。
想要治理天下,却又要用强制的办法,我看他不能够达到目的。
天下神器,不可为也。不可执也
天下的人民是神圣的,不能够违背他们的意愿和本性而加以强力统治,不能横加干涉去左右和控制制约强力把持
为者败之,执者失之。
用强力纺治天下,就一定会失败;强力把持天下,就一定会失去
是以圣人无为,故无败,故无失
。因此,圣人不妄为,所以不会失败;不把持,所以不会被抛弃。
故物或行或随;或嘘或吹,或强或羸,或挫或隳 。
[ huī ] 毁坏、损毁。 【组词】:隳坏 [ duò ] 古通“惰”,懒惰。
是以圣人去甚,去奢,去泰。
因此,圣人要除去那种极端、奢侈的、过度的措施法度。
世人秉性不一,有前行有后随,有轻嘘有急吹,有的刚强,有的赢弱;有的安居,有的危殆。
以道佐人主者,不以兵强天下。
依照“道”的原则辅佐君主的人,不以兵力逞强于天下。
其事好远。
穷兵黩武这种事必然会得到报应
果而勿矜,果而勿伐,果而勿骄。
果而勿矜,
达到目的了却不自我矜持
果而勿伐
达到目的了也不去夸耀骄傲
果而勿骄
达到目的了也不要自以为是
师之所处,荆棘生焉;大军之后,必有凶年。
军队所到的地方,荆棘横生,大战之后,一定会出现荒年。
果而不得已,果而勿强。
果而不得已,
,达到目的却出于不得已,
果而勿强。
达到目的却不逞强。
善有果而已,不敢以取强。
善于用兵的人,只要达到用兵的目的也就可以了,并不以兵力强大而逞强好斗。
道常无名,朴虽小,天下莫能臣。
“道”永远是无名而质朴的,它虽然很小不可见,天下没有谁能使它服从自己
侯王若能守之,万物将自宾。
如果能够依照“道”的原则治理天下,百姓们将会自然地归从于它。
天地相合,以降甘露,民莫之令而自均。
天地间阴阳之气相合,就会降下甘露,人们不必指使它而会自然均匀。
始制有名,名亦既有,夫亦将知止。
治理天下就要建立一种管理体制,制定各种制度确定各种名分,任命各级官长办事。名分既然有了,就要有所制约,适可而止,
知止可以不殆。
知道制约、适可而止,就没有什么危险了。
譬道之在天下,犹川谷之于江海。
“道”存在于天下,就像江海,一切河川溪水都归流于它,使万物自然宾服。
知人者智,自知者明;胜人者有力,自胜者强;知足者富,强行者有志;不失其所者久,死而不亡者寿。
知人者智,
能了解、认识别人叫做智慧,
自知者明;
能认识、了解自己才算聪明。
胜人者有力
能战胜别人是有力的
自胜者强
能克制自己的弱点才算刚强。
知足者富
知道满足的人才是富有人
强行者有志;
坚持力行、努力不懈的就是有志。
不失其所者久,
离失本分的人就能长久不衰,
死而不亡者寿。
身虽死而“道”仍存的,才算真正的长寿。
将欲歙之,必固张之;将欲弱之,必固强之;将欲废之,必固举之;将欲取之,必固予之。
将欲歙之,必固张之;
想要收敛它,必先扩张它
xī 歙:吸气或通过呼吸吸入
将欲弱之,必固强之;
想要削弱它,必先加强它,
将欲废之,必固举之;
想要废去它,必先抬举它,
将欲取之,必固予之。
想要夺取它,必先给予它
鱼不可脱于渊,国之利器不可以示人。
鱼的生存不可以脱离池渊,国家的刑法政教不可以向人炫耀,不能轻易用来吓唬人。
道常无为,而无不为。
道永远是顺任自然而无所作为的,却又没有什么事情不是它所作为的。
侯王若能守之,万物将自化。
侯王如果能按照“道”的原则为政治民,万事万物就会自我化育、自生自灭而得以充分发展。
化而欲作,吾将镇之以无名之朴。
自生自长而产生贪欲时,我就要用“道”来镇住它
不欲以静,天下将自定。
万事万物没有贪欲之心了,天下便自然而然达到稳定、安宁。
修德
修德
高贵必然要以贫贱拥戴为根本,高处必然要以下面的支撑为基础
最高的荣誉无须赞美称誉
不要求琭琭晶莹像宝玉,而宁愿珞珞坚硬像山石
应该立身敦厚,存心朴实,不居浇薄,不居虚化
浇薄,汉语词汇,拼音为jiāo bó,意思是薄,不厚,贫薄,指社会风气浮薄。
总结
A、道生一,一生二,二生三,三生万物。
B、万物负阴而抱阳,冲气以为和
重点注意
一切事物,如果减损它却反而得到增加;如果增加它却反而得到减损
要注意事情的辩证处理方式对本人而言
强暴的人死无其所
用强会有很大的阻力,很多都不会成功
不言之教,无为之益,天下希及之。
“不言”的教导,“无为”的益下,普天下少有能赶上它的了。
甚爱必大费;厚藏必多亡。
过分的爱名利就必定要付出更多的代价;过于积敛财富,必定会遭致更为惨重的损失
故知足不辱,知止不殆,可以长久。
所以说,懂得满足,就不会受到屈辱;懂得适可而止,就不会遇见危险;这样才可以保持住长久的平安。
大直若屈,大巧若拙,大辩若讷。
最正直的东西,好似有弯曲一样;最灵巧的东西,好似最笨拙的;最卓越的辩才,好似不善言辞一样
核心
清静无为才能统治天下
不妄为
顺势
取天下常以无事,及其有事,不足以取天下。
治理国家的人,要经常以不骚扰人民为治国之本,如果经常以繁苛之政扰害民众,那就不配治理国家了。
无为而无不为。
如果能够做到无为,即不妄为,任何事情都可以有所作为。
为学日益,为道日损。
求学的人,其情欲文饰一天比一天增加;求道的人,其情欲文饰则一天比一天减少
圣人之在天下,歙歙焉,为天下浑其心,百姓皆注其耳目焉,圣人皆咳之。
有道的圣人在其位,收敛自己的欲意,使天下的心思归于浑朴。百姓们都专注于自己的耳目聪明,有道的人使他们都回到婴孩般纯朴的状态。
圣人恒无心,以百姓之心为心。
圣人常常是没有私心的,以百姓的心为自己的心
盖闻善摄生者,陆行不避兕虎,入军不被甲兵;兕无所投其角,虎无所用其爪,兵无所容其刃。
盖闻善摄生者,陆行不避兕虎,入军不被甲兵;
据说,善于养护自己生命的人,在陆地上行走,不会遇到凶恶的犀牛和猛虎,在战争中也受不到武器的伤害
(sì)兕
上古瑞兽“兕”,状如犀牛不是犀牛(雷兽科),苍黑(全身呈现青黑色),板角(就是独角兽那样的犄角)。逢天下将盛,而现世出。 “兕”中最著名的是板角青牛(太上老君的坐骑) 古代指犀牛(一说雌性犀牛)
兕无所投其角,虎无所用其爪,兵无所容其刃。
犀牛于其身无处投角,老虎对其身无处伸爪,武器对其身无处刺击锋刃
以其无死地焉。
因为他没有进入死亡的领域。
万物本身都有起始,这个作为天地万物的根源,知其根源就能认识万物,认识万物又把握万物的根源,终身没有危险,要见微知著,持守柔弱乃强
走平坦大道,不走邪路
总结
A、把这个道理付诸自身,他的德行就会真实纯真,把这个道理付诸于自家,他的德性就会是丰盈有余的;把这个道理付诸于自乡,他的德性就会受到尊崇;把这个道理付诸于自邦,他的德性就会丰盛硕大;把这个道理付诸于天下,他的德性就会无限普及。
1、天地万物本身都有起始,这个始作为天地万物的根源
2、知道根源,认识万物
3、认识了万事万物,又把握着万物的根本
4、塞住欲念的孔穴,闭起欲念的门径,
5、能够察见到细微的;能够持守柔弱的。
6、运用其(德)光芒,返照内在的明,遵循“常道”
7、养育万物,使万物生长发展,成熟结果,使其受到抚养、保护
8、生长万物而不居为己有,抚育万物而不自恃有功,导引万物而不主宰
9、不奉养太过度
10、不进入死亡的领域
善于养护自己生命的人,在陆地上行走,不会遇到凶恶的犀牛和猛虎,在战争中也受不到武器的伤害
犀牛于其身无处投角,老虎对其身无处伸爪,武器对其身无处刺击锋刃
11、能够做到无为,即不妄为,任何事情都可以有所作为。
12、知道到什么地步就该满足
13、道是独一无二的,道本身包含阴阳二气,阴阳二气相交而形成一种适匀的状态,万物在这种状态中产生。
14、行“不言”的教导,清净“无为”。
15、具备“上德”不表现为外在的有德,因此实际上是有“德”;
B、用自身的修身之道来观察别身;以自家之道察看观照别家;以自乡之道察看观照别乡;以平天下之道察看观照天下。
道德涵养浑厚的人
就好比初生的婴孩
毒虫不螫他,猛兽不伤害他,凶恶的鸟不搏击他
他的筋骨柔弱,但拳头却握得很牢固
他虽然不知道男女的交合之事,但他的小生殖器却勃然举起,这是因为精气充沛的缘故
他整天啼哭,但嗓子却不会沙哑,这是因为和气纯厚的缘故
天下人尊重的几个重要的点
1、知者不言,言者不知
2、管住嘴,不露锋芒,消解纷争,收敛光辉,混同尘世
3、超脱亲疏,利害,贵贱的世俗范围
其政闷闷,其民淳淳;其政察察,其民缺缺。
政治宽厚清明,人民就淳朴忠诚;政治苛酷黑暗,人民就狡黠、抱怨
是以圣人方而不割,廉而不刿,直而不肆,光而不耀。
。因此,有道的圣人方正而不生硬,有棱角而不伤害人,直率而不放肆,光亮而不刺眼。
治理百姓养护身心,最重要的是爱惜精神力
爱惜精神,得以早做准备
早做准备,可以不断的积德
不断地积德,就没有什么不能攻克的
攻无不克,就没有办法估量他的力量
具备无法估量的力量就可以治理国家
有了整理国家的原则和道理,国家就可以长久
遵循道以德治理天下更为合理
鬼神起不了作用,鬼怪的作用也伤不了人,圣人有道也伤不了人
圣人和鬼神都不伤害,就可以然人民享受德的恩泽
美言可以市,尊行可以加人。
美好的言辞可以换来别人对你的尊重;良好的行为可以见重于人
重要的守则
以无为的态度有所作为,以不滋事方法处理事务,以恬淡无味当作有味
处理问题要从容易的地方入手,实现远大要从细微的地方入手
天下难事,必作于易;天下大事,必作于细。
那些轻易发出诺言的,必定很少能够兑现的,把事情看得太容易,势必遭受很多困难
重要守则
其安易持,其未兆易谋。
局面安定时容易保持和维护,事变没有出现迹象时容易图谋;
其脆易泮,其微易散。
事物脆弱时容易消解;事物细微时容易散失
为之于未有,治之于未乱。
做事情要在它尚未发生以前就处理妥当;治理国政,要在祸乱没有产生以前就早做准备
合抱之木,生于毫末;九层之台,起于累土;千里之行,始于足下。
合抱的大树,生长于细小的萌芽;九层的高台,筑起于每一堆泥土;千里的远行,是从脚下第一步开始走出来的。
为者败之,执者失之。
有所作为的将会招致失败,有所执着的将会遭受损害。
妄为会招致失败
是以圣人无为故无败,无执故无失。
因此圣人无所作为所以也不会招致失败,无所执着所以也不遭受损害
民之从事,常于几成而败之。
人们做事情,总是在快要成功时失败
不慎终也。慎终如始,则无败事。
所以当事情快要完成的时候,也要像开始时那样慎重,就没有办不成的事情
是以圣人欲不欲,不贵难得之货;学不学,复众人之所过。
因此,有道的圣人追求人所不追求的,不稀罕难以得到的货物,学习别人所不学习的,补救众人所经常犯的过错。
以辅万物之自然而不敢为。
这样遵循万物的自然本性而不会妄加干预。
玄德深矣远矣,与物反矣,然后乃至大顺。
玄德又深又远,和具体的事物复归到真朴,然后才能极大地顺乎于自然。
古代善于为道的人,不是教导人民知晓智巧伪诈,而是教导人民淳厚朴实
民之难治,以其智多。
人们之所以难于统治,乃是因为他们使用太多的智巧心机。
故以智治国,国之贼;不以智治国,国之福。
所以用智巧心机治理国家,就必然会危害国家,不用智巧心机治理国家,才是国家的幸福。
江海所以能为百谷王者,以其善下之,故能为百谷王。
江海所以能够成为百川河流所汇往的地方,乃是由于它善于处在低下的地方,所以能够成为百川之王
是以欲上民,必以言下之;欲先民,必以身后之。
因此,圣人要领导人民,必须用言辞对人民表示谦下,要想领导人民,必须把自己的利益放在他们的后面。
是以圣人处上而民不重,处前而民不害。
所以,有道的圣人虽然地位居于人民之上,而人民并不感到负担沉重,居于人民之前,而人民并不感到受害。
是以天下乐推而不厌,以其不争,故天下莫能与之争。
天下的人民都乐意推戴而不感到厌倦。因为他不与人民相争,所以天下没有人能和他相争。
一曰慈,二曰俭,三曰不敢为天下先。
第一件叫做慈爱;第二件叫做俭啬;第三件是不敢居于天下人的前面。
慈故能勇;俭故能广;不敢为天下先,故能成器长。
有了这柔慈,所以能勇武;有了俭啬,所以能大方;不敢居于天下人之先,所以能成为万物的首长。
善为士者不武,善战者不怒,善胜敌者弗与;善用人者,为之下。
善为士者不武,
善于带兵打仗的将帅,不逞其勇武;
善战者不怒
善于打仗的人,不轻易激怒;
善胜敌者弗与
善于胜敌的人,不与敌人正面冲突
善用人者,为之下
善于用人的人,对人表示谦下
是谓不争之德,是谓用人之力,是谓配天古之极。
这叫做不与人争的品德,这叫做运用别人的能力,这叫做符合自然的道理。
是谓行无行,攘无臂,执无兵,乃无敌矣;祸莫大于轻敌,轻敌几丧吾宝。
是谓行无行
这就叫做虽然有阵势,却像没有阵势可摆一样
攘无臂
虽然要奋臂,却像没有臂膀可举一样
扔无敌
虽然面临敌人,却像没有敌人可打一样
执无兵
虽然有兵器,却像没有兵器可以执握一样
乃无敌矣
所以无敌
祸莫大于轻敌,轻敌几丧吾宝
祸患再没有比轻敌更大的了,轻敌几乎丧失了我的“三宝”。
第一件叫做慈爱;第二件叫做俭啬;第三件是不敢居于天下人的前面。(三宝)
吾言甚易知,甚易行。
我的话很容易理解,很容易施行
天下莫能知,莫能行。
但是天下竟没有谁能理解,没有谁能实行
言有宗,事有君。
言论有主旨,行事有根据
夫唯无知,是以不我知。
正由于人们不理解这个道理,因此才不理解我
知我贵希,是以圣人被褐而怀玉。
因此有道的圣人总是穿着粗布衣服,怀里揣着美玉。
知不知,尚矣;不知知,病也;是以圣人不病也,是以不病。
知不知,尚矣;
知道自己还有所不知,这是很高明的
不知知,病也;
不知道却自以为知道,这就是很糟糕的
是以圣人不病也,是以不病
有道的圣人没有缺点,因为他把缺点当作缺点。正因为他把缺点当作缺点,所以,他没有缺点。
民不畏威,则大威至;无狎其所居,无厌其所生。
民不畏威,则大威至;
当人民不畏惧统治者的威压时,那么,可怕的祸乱就要到来了
无狎其所居,无厌其所生
不要逼迫人民不得安居,不要阻塞人民谋生的道路。
夫唯不厌,是以不厌。
只有不压迫人民,人民才不厌恶统治者
是以圣人不病也,是以不病
有道的圣人没有缺点,因为他把缺点当作缺点。正因为他把缺点当作缺点,所以,他没有缺点。
天之道,不争而善胜,不言而善应,不召而自来,坦然而善谋。
天之道
不争而善胜
不斗争而善于取胜
不言而善应
不言语而善于应承
不召而自来
不召唤而自动到来
坦然而善谋
坦然而善于安排筹划
自然的规律是
天网恢恢,疏而不失
自然的范围,宽广无边,虽然宽疏但并不漏失
重要守则
若民恒且不畏死,奈何以杀惧之也?
人民不畏惧死亡,为什么用死来吓唬他们呢?
若民恒且畏死,则为奇者,吾将得而杀之,夫孰敢矣?
假如人民真的畏惧死亡的话,对于为非作歹的人,我们就把他抓来杀掉。谁还敢为非作歹?
若民恒且必畏死,则恒有司杀者夫代司杀者杀,是代大匠斫也,夫代大匠斫者,希有不伤其手矣。
若民恒且必畏死,则恒有司杀者
假如百姓历来害怕死亡,那么应该历来就有专门负责杀戮的机构
夫代司杀者杀,是代大匠斫[zhuó]也
如果代替掌管杀戮者去杀戮,就如同代替高超的木匠去砍削木头
夫代大匠斫者,则希不伤其手矣。
而代替高超的木匠砍削木头的人,则很少能有不砍伤自己手指头的
民之难治,以其上之有为,是以难治;
人民之所以难于统治,是由于统治者政令繁苛、喜欢有所作为,所以人民就难于统治
民之轻死,以其求生之厚,是以轻死。
人民之所以轻生冒死,是由于统治者为了奉养自己,把民脂民膏都搜刮净了,所以人民觉得死了不算什么
天之道,损有余而补不足。
世人现在的了解是物竞天择适者生存,所以应该是损不足而补有余?但是和这个并不冲突
自然的规律,是减少有余的补给不足的
人之道则不然,损不足以奉有余。
可是社会的法则却不是这样,要减少不足的,来奉献给有余的人
孰能有余以奉天下,唯有道者。
那么,谁能够减少有余的,以补给天下人的不足呢?只有有道的人才可以做到。
是以圣人为而不恃,功成而不处,其不欲见贤邪。
因此,有道的圣人这才有所作为而不占有,有所成就而不居功。他是不愿意显示自己的贤能。
是以圣人云:受国之垢,是谓社稷主;受国不祥,是为天下王。
所以有道的圣人这样说:“承担全国的屈辱,才能成为国家的君主,承担全国的祸灾,才能成为天下的君王。”
是以圣人执左契,而不责于人。
因此,有道的圣人保存借据的存根,但并不以此强迫别人偿还债务。
有德司契,无德司彻。
有“德”之人就像持有借据的圣人那样宽容,没有“德”的人就像掌管税收的人那样苛刻刁诈。
夫天道无亲,常与善人。
自然规律对任何人都没有偏爱,永远帮助有德的善人
重要守则
信言不美,美言不信。
真实可信的话不漂亮,漂亮的话不真实
善者不辩,辩者不善。
善良的人不巧说,巧说的人不善良
知者不博,博者不知。
真正有知识的人不卖弄,卖弄自己懂得多的人不是真有知识
圣人不积:既以为人己愈有;既以与人,己愈多。
圣人不积:既以为人己愈有;
圣人是不存占有之心的,而是尽力照顾别人,他自己也更为充足;
既以与人,己愈多。
他尽力给予别人,自己反而更丰富
天之道,利而不害;人之道,为而弗争。
天之道,利而不害;
自然的规律是让万事万物都得到好处,而不伤害它们
人之道,为而弗争。
圣人的行为准则是,做什么事都不跟别人争夺。