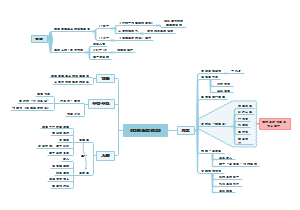
对脑干和间脑促眠区施以低频电刺激可引起NREM睡眠,而施以高频电刺激则引起觉醒;而在前脑促眠区无论施加低频或高频刺激均将引起NREM睡眠的发生。
脑内存在多个促进NREM睡眠的部位,其中最重要的是视前区腹外侧部( VLPO)。
由觉醒进入NREM睡眠后,VLPO神经元放电频率增高,且细胞原癌基因c-fos表达增加(表示此时处于活动状态)。VLPO内存在大量促睡眠神经元,它们发出的纤维投射到脑内多个与觉醒有关的部位,如蓝斑去甲肾上腺素能神经元、中缝背核5-羟色胺能神经元脑桥头端被盖胆碱能神经元、下丘脑结节乳头体核组胺能神经元等,VLPO投射纤维的主要递质是y-氨基丁酸,通过对促觉醒脑区活动的抑制,促进觉醒向睡眠转化,产生NREM睡眠。
有研究表明,视交叉上核有纤维通过其他核团中继后投射到下丘脑外侧部的增食因子能神经元和VLPO,将昼夜节律的信息传递给促觉醒和促睡眠脑区,调节觉醒与睡眠的相互转换。
促进NREM睡眠的脑区还位于延髓网状结构的脑干促眠区(也称上行抑制系统)
位于下丘脑后部、丘脑髓板内核群邻旁区和丘脑前核的间脑促眠区
位于下丘脑或前脑视前区和Broca斜带区的前脑基底部促眠区