导图社区 神州谣
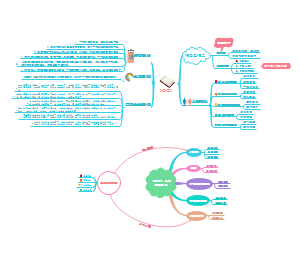
神州谣
小学二年级下册语文部编版神州谣思维导图
编辑于2020-03-26 15:01:35- 鲁迅朝花夕拾思维导图
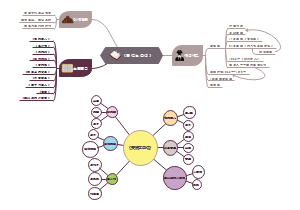
鲁迅朝花夕拾思维导图,作者渐渐、作品介绍、主题思想、十个故事的意义。《朝花夕拾》共收入10篇作品,这些作品以青少年时代的生活经历为线索,真实生动地叙写了鲁迅从农村到城镇,从家庭到社会,从国内到国外的一组生活经历。
- 小学、初中语文阅读理解拟人全解
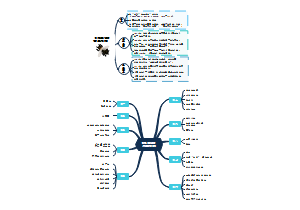
小学初中 阅读理解 拟人部分 比喻与拟人的区别 如何答题,介绍了比喻句的分类,包括明喻、借喻和暗喻,并强调了比喻句中喻体的重要性。通过具体的例子说明了拟人句的特点,即没有喻体,而是将事物进行人格化的描述,赋予其人的情感和行为。还详细解释了拟人句如何通过模仿人的行为或情感来生动形象地描述事物,并通过多个例子加以说明。展示了比喻修辞的寻找喻体和比较喻体的过程,以及比喻和拟人的比较。通过具体的练习,读者可以加深对这两种修辞手法的理解和应用。
- 小学、初中语文根据上下文解释词义
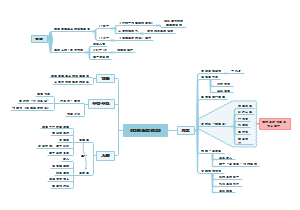
根据上下文解释词义(解决七字诀),本指喻指文中指,答题方法,习题,,“本指”是词语最基础、最原本的含义,可以通过拆词法、近义词法和否定反义词法来理解。例如,“寸草不生”通过拆词法被拆解为“寸”、“草”和“不生”,再组合成新的含义“一寸小草也没有生长”。近义词法则是通过替换词语的方式来解释词义,如“郁郁葱葱”可以换成“枝繁叶茂”。否定反义词法则是在词语前加上否定词并找出其反义词来形成新的解释等内容,结构清晰,易于阅读和理解。此外,还提供了习题助记部分,通过实例来加深理解和记忆。为理解和记忆中文词语提供了一种系统化和多样化的方法。
神州谣
社区模板帮助中心,点此进入>>
- 鲁迅朝花夕拾思维导图
鲁迅朝花夕拾思维导图,作者渐渐、作品介绍、主题思想、十个故事的意义。《朝花夕拾》共收入10篇作品,这些作品以青少年时代的生活经历为线索,真实生动地叙写了鲁迅从农村到城镇,从家庭到社会,从国内到国外的一组生活经历。
- 小学、初中语文阅读理解拟人全解
小学初中 阅读理解 拟人部分 比喻与拟人的区别 如何答题,介绍了比喻句的分类,包括明喻、借喻和暗喻,并强调了比喻句中喻体的重要性。通过具体的例子说明了拟人句的特点,即没有喻体,而是将事物进行人格化的描述,赋予其人的情感和行为。还详细解释了拟人句如何通过模仿人的行为或情感来生动形象地描述事物,并通过多个例子加以说明。展示了比喻修辞的寻找喻体和比较喻体的过程,以及比喻和拟人的比较。通过具体的练习,读者可以加深对这两种修辞手法的理解和应用。
- 小学、初中语文根据上下文解释词义
根据上下文解释词义(解决七字诀),本指喻指文中指,答题方法,习题,,“本指”是词语最基础、最原本的含义,可以通过拆词法、近义词法和否定反义词法来理解。例如,“寸草不生”通过拆词法被拆解为“寸”、“草”和“不生”,再组合成新的含义“一寸小草也没有生长”。近义词法则是通过替换词语的方式来解释词义,如“郁郁葱葱”可以换成“枝繁叶茂”。否定反义词法则是在词语前加上否定词并找出其反义词来形成新的解释等内容,结构清晰,易于阅读和理解。此外,还提供了习题助记部分,通过实例来加深理解和记忆。为理解和记忆中文词语提供了一种系统化和多样化的方法。
- 相似推荐
- 大纲