导图社区 炎症相关的硝酸盐促进口腔V. parvula在肠道内异位定植
炎症相关的硝酸盐促进口腔V. parvula在肠道内异位定植
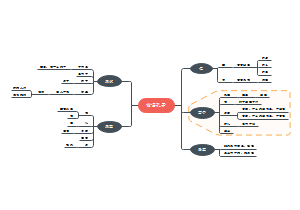
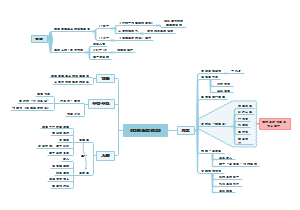
炎症相关的硝酸盐促进口腔V. parvula在肠道内异位定植:肠道炎症与V.parvula富集有关、硝酸盐呼吸作用扩展了V.parvula的代谢范围、硝酸盐呼吸作用允许V.parvula利用氨基酸生长……
编辑于2022-10-04 21:49:07 湖北省- parvula利用氨基酸生长
- 炎症相关的硝酸盐促进口腔V. parvula在肠道内异位定植
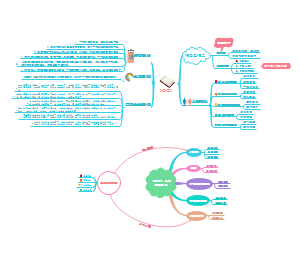
炎症相关的硝酸盐促进口腔V. parvula在肠道内异位定植,包括:口腔厌氧菌韦荣球菌属物种在炎症性肠病患者肠道中丰度升高、韦荣球菌能利用炎症标志性代谢产物硝酸盐,依赖narGHJI操纵子的硝酸盐呼吸依赖narGHJI操纵子的硝酸盐呼吸,可促进韦荣球菌属在有机酸中的生长、硝酸盐呼吸可促进韦荣球菌属利用氨基酸和肽作为碳源,这种代谢转变伴随着碳代谢的变化、硝酸盐呼吸可促进韦荣球菌属利用氨基酸和肽作为碳源,这种代谢转变伴随着ATP合成途径的变化。
- 炎症相关的硝酸盐促进口腔V. parvula在肠道内异位定植
炎症相关的硝酸盐促进口腔V. parvula在肠道内异位定植:肠道炎症与V.parvula富集有关、硝酸盐呼吸作用扩展了V.parvula的代谢范围、硝酸盐呼吸作用允许V.parvula利用氨基酸生长……
- 酪酸梭菌通过 TLR2MyD88NF-κB 信号通路调控
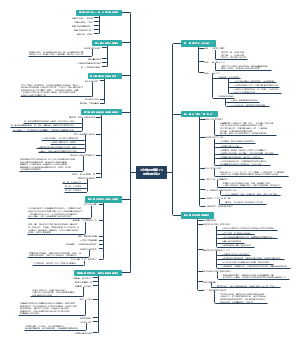
酪酸梭菌通过 TLR2MyD88NF-κB 信号通路调控:酪酸梭菌通过调控 TLR2/MyD88/NF-κB 信号通路途径调节 DSS 诱导的实验性结肠炎小鼠异常免疫反应的机制研究。
炎症相关的硝酸盐促进口腔V. parvula在肠道内异位定植
社区模板帮助中心,点此进入>>
- 炎症相关的硝酸盐促进口腔V. parvula在肠道内异位定植
炎症相关的硝酸盐促进口腔V. parvula在肠道内异位定植,包括:口腔厌氧菌韦荣球菌属物种在炎症性肠病患者肠道中丰度升高、韦荣球菌能利用炎症标志性代谢产物硝酸盐,依赖narGHJI操纵子的硝酸盐呼吸依赖narGHJI操纵子的硝酸盐呼吸,可促进韦荣球菌属在有机酸中的生长、硝酸盐呼吸可促进韦荣球菌属利用氨基酸和肽作为碳源,这种代谢转变伴随着碳代谢的变化、硝酸盐呼吸可促进韦荣球菌属利用氨基酸和肽作为碳源,这种代谢转变伴随着ATP合成途径的变化。
- 炎症相关的硝酸盐促进口腔V. parvula在肠道内异位定植
炎症相关的硝酸盐促进口腔V. parvula在肠道内异位定植:肠道炎症与V.parvula富集有关、硝酸盐呼吸作用扩展了V.parvula的代谢范围、硝酸盐呼吸作用允许V.parvula利用氨基酸生长……
- 酪酸梭菌通过 TLR2MyD88NF-κB 信号通路调控
酪酸梭菌通过 TLR2MyD88NF-κB 信号通路调控:酪酸梭菌通过调控 TLR2/MyD88/NF-κB 信号通路途径调节 DSS 诱导的实验性结肠炎小鼠异常免疫反应的机制研究。
- 相似推荐
- 大纲