导图社区 细胞内遗传信息的传递及调控
本图是对细胞生物学之细胞内遗传信息的传递及调控的知识梳理,内容涵盖人类对基因的认识和探索过程、基因及其结构、基因转录和转录后加工三部分,本专业和有兴趣的朋友们不要错过。
细胞核(nucleus)是真核细胞内最大、最重要的细胞结构,是细胞遗传与代谢的调控中心,是真核细胞区别于原核细胞最显著的标志之一(极少数真核细胞无细胞核,如哺乳动物的成熟的红细胞,高等植物成熟的筛管细胞等)
医学细胞生物学,详细的总结了细胞骨架。微管,中间纤维,微丝的内容点。帮助小伙伴快速掌握医学细胞生物学的内容要点!
社区模板帮助中心,点此进入>>
论语孔子简单思维导图
《傅雷家书》思维导图
《童年》读书笔记
《茶馆》思维导图
《朝花夕拾》篇目思维导图
《昆虫记》思维导图
《安徒生童话》思维导图
《鲁滨逊漂流记》读书笔记
《这样读书就够了》读书笔记
妈妈必读:一张0-1岁孩子认知发展的精确时间表
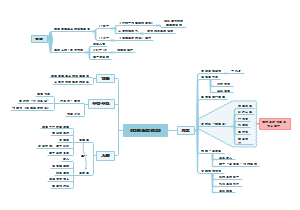
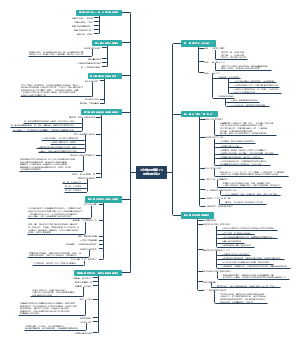
细胞内遗传信息的传递及调控
人类对基因的认识和探索过程
J.G.Mendel 奥地利 1854年
分离定律和自由组合定律
一个因子决定一个性状
A. Weismann 德国 1892年
种质论主张生物体由质上根本相异的两部分——种质和体质组成
生物体在一生中由于外界环境的影响或器官的用与不用所造成的变化只表现于体质上,而与种质无关,所以后天获得性状不能遗传
种质只存在于核内染色质中
T.H.Morgan 美国 1926年
基因学说
基因是组成染色体的遗传单位,它能控制遗传性状的发育,也是突变、重组、交换的基本单位
J.Watson&F.Crick 美国 英国 1953年
DNA双螺旋模型
F.Crick 1958年
中心法则
基因及其结构
基因是DNA分子中含有特定遗传信息的核苷酸序列
基因
细胞内遗传物质的最小功能单位,是负载有特定遗传信息的DNA片段
结构基因
编码非调控因子的任何蛋白质和RNA的基因(结构蛋白、酶、rRNA和tRNA)
调控基因
通过编码蛋白或RNA来调节其他基因的表达
基因组
指细胞或生物体的一套完整的单倍体遗传物质,是所有染色体上全部基因和基因间的DNA的总和,它含有一个生物体进行各种生命活动所需要的全部遗传信息
人类基因组约有3.0×10⁹bp,2~3万个基因
中心法则揭示了基因的信息流向
遗传信息从DNA到RNA再到蛋白质的流动,称为分子生物学的中心法则
包括
DNA复制
遗传信息由亲代DNA分子传给子代DNA
RNA转录
以DNA为模板合成RNA
蛋白质翻译
RNA(mRNA)指导合成蛋白质的过程
补充
RNA→DNA
原核细胞的基因结构
大多数原核细胞中只有一个DNA分子,原核细胞基因组DNA的绝大多数可以编码蛋白质,少部分不转录
编码区
能够转录为相应的mRNA,进而指导蛋白质的基因区段
在原核细胞中是连续的,功能相关的结构基因串联排列
编码蛋白质的区段由编码区上游和下游的DNA序列组成
启动子
位于结构基因上游,包含转录起始点和RNA聚合酶识别部位和结合部位
起始点
DNA模板链上开始进行转录作用的位点
识别部位
RNA聚合酶的σ因子识别DNA分子的部位,中心位于-35区
共有序列是5'-TTGACA-3'
结合部位
与RNA聚合酶核心酶紧密结合的序列,中心位于-10区
共同序列为5'-TATAAT-3',又称为Pribnow盒
非编码区
占整个基因组小部分,不能转录为mRNA
真核细胞的基因结构
基因由多个功能区域组成
由编码区和非编码区两部分组成,编码区是不连续的,被非编码区所隔断,因而真核细胞基因也被称为断裂基因
完整的真核基因包括外显子、内含子、启动子和终止子
外显子
指在结构基因中能够被转录,并能指导蛋白质生物合成的编码序列
内含子
指插入在结构基因内部能够被转录,但不能指导蛋白质生物合成的非编码序列
真核细胞启动子由TATA盒及其上游的CAAT盒和/或GC盒组成
TATA box
在转录起点-25~-35区段由TATAATAAT组成的序列
是RNA聚合酶结合位点,与转录起始的准确定位有关
CAAT box
在转录起点-70~-80区段由CCAAT组成的序列
是蛋白质转录因子的结合位点,控制转录的起始频率
GC box
在转录起点-80~-110区段由GGGCGG组成的序列
是蛋白质转录因子的识别位点,控制转录的起始频率
终止子
存在于基因末端具有转录终止功能的特定序列
转录后形成发夹结构,使RNA聚合酶从模板上脱离,终止转录
存在多个基因家族
基因家族
真核细胞基因组中来源相同、结构相似、功能相关的一组基因,是由一个祖先基因经重复和变异形成的,是真核细胞基因结构最显著的特征之一
家族成员的存在形式
成簇存在
串联排列于特殊的染色体区段上,形成基因簇
可同时转录、合成功能相关或相同的产物;如组蛋白、rRNA基因家族
分散存在
广泛分布于整个染色体,甚至可存在于不同的染色体上
如干扰素、珠蛋白等基因家族
含有大量的DNA重复序列
基因转录和转录后加工
基因转录的一般特点
基因转录
是遗传信息从DNA流向RNA的过程,即将DNA分子上的核苷酸序列转变为RNA分子上核苷酸序列的过程
转录的本质
以DNA双螺旋中的一条链为模板
以四种核苷三磷酸ATP、GTP、CTP、UTP为原料
在RNA聚合酶作用下,遵循碱基互补配对原则,合成RNA的过程
模板链
又称或反义链,在双链DNA中,作为转录模板的链,与mRNA互补
编码链
又称或有意义链,在双链DNA中,与模板链互补的另一条链,与转录产物的序列相同
不对称转录
DNA双链的某一区段,以其中一条单链为模板,而在另一区段,以其相对应的互补链为模板,这种DNA链的选择性转录称为不对称转录
蛋白质的生物合成
翻译
细胞以mRNA作为模板,tRNA作为运输工具,rRNA和蛋白质组成的核糖体作为装配机器,合成蛋白质的过程
mRNA携带指导蛋白质合成的遗传密码
遗传密码
mRNA上的碱基排列顺序
特点
通用性
从原核生物到真核生物,几乎所有生物体中的遗传密码都是通用的
简并性
多个密码子决定同一氨基酸
连续性
在翻译过程中,遗传密码的阅读都是连续的
方向性
在mRNA中密码子的阅读方向为5'→3'
密码子
在mRNA链上3个相邻的碱基可以决定一个特定的氨基酸,这种核苷酸三联体被称为密码子
64组密码子,20种氨基酸
遗传信息翻译的基本原理
转运RNA
tRNA识别mRNA上的密码子
反密码子的碱基互补配对作用
tRNA携带特定氨基酸
氨酰-tRNA合成酶催化形成氨酰-tRNA
配对的摆动性
密码子的前两位碱基在和反密码子配对时,遵循正常的碱基互补配对原则,而第三位碱基的配对具有一定的灵活性
核糖体
核糖体是一种非膜相结构的颗粒状细胞器,由核糖核酸和蛋白质组成,又称核糖核酸蛋白质
核糖体是细胞内蛋白质生物合成的场所
发现历史
1953年—Robinson&Brown—植物细胞 1955年—Palade—电镜—腺细胞
主要成分
核糖体蛋白(r蛋白质) 占总量的1/3
核糖核酸 占总量的2/3
rRNA
tRNA
主要分布
核糖体常分布在细胞内蛋白质合成旺盛的区域
形态结构
无生物膜包被
直径25~30nm致密椭圆形小颗粒状的结构
由大、小亚基两个部分组成
大亚基约为小亚基的两倍
大亚基
大亚基略呈圆锥形,侧面观其两侧隆起,底边为扁平状,有一条很窄的沟
底面能与小亚基相接,中央有一中央管,为新合成的多肽链释放的通道
小亚基
呈略微弯曲的葫芦形
一面略外凸,一面略凹陷,中段有一分界线,将其分成两个不等的部分
完整的核糖体中,小亚基凹面与大亚基的扁平部相贴
小亚基中间分界线与大亚基部的沟相吻合,在大、小亚基结合面上形成一条隧道,是mRNA穿过的通道。
中央有一中央管,为新合成的多肽链释放的通道
基本类型
原核生物核糖体
胞质核糖体
相对分子量2.5×10⁶,沉降系数70S
真核生物核糖体
植物
细胞质核糖体
线粒体核糖体
叶绿体核糖体
动物
相对分子量4.2×10⁶,沉降系数80S
核糖体结构位点
mRNA结合位点
原核
SD序列,与小亚基16S rRNA互补结合
真核
5'-甲基化帽子
tRNA结合位点
A位点
氨酰基位点
P位点
肽酰基位点
E位点
tRNA释放位点
转移酶活性部位
催化氨基酸形成肽链
蛋白合成因子结合部位
结合起始因子
延长因子
释放因子
多聚核糖体
具有特殊形态结构的核糖体与mRNA的聚合体
多核糖体利用同一条mRNA同时进行蛋白质合成,大大提高蛋白质合成的效率
一般过程
预备阶段:氨基酸活化
起始阶段:起始复合物的装配
30S小亚基与mRNA结合
起始氨酰-tRNA进入核糖体
完整起始复合物的装配
延伸阶段:新合成肽链的成长
氨酰-tRNA进入核糖体A位点
肽键的形成
A位点氨基酸的氨基与P位点氨基酸的羧基,在肽酰转移酶的作用下,形成肽键
转位
核糖体沿着mRNA5'-3'方向移动3个核苷酸
脱氨酰-tRNA的释放
终止阶段:肽链合成的终止
多肽链的释放,大小亚基解体,mRNA降解
加工阶段:肽链合成后的加工修饰
新生多肽链不具有生物活性,需经过化学修饰和加工处理才能形成具有天然构象和生物学活性的功能蛋白
(一)一级结构的修饰改变了多肽链氨基酸的性质和组成
肽链氨基端的修饰
N-甲酰甲硫氨酸或甲硫氨酸的去除:甲酰基经脱甲酰基酶水解,甲硫氨酸或者氨基酸的一些氨基酸残基由氨肽酶水解
共价修饰
磷酸化、糖基化、羟基化、甲基化、乙酰化和二硫键的形成等
多肽链的水解修饰
前体蛋白水解产生活性蛋白
多肽链经水解后产生几种不同的蛋白质或多肽
浮动主题
cover1782_20221122081501.jpg?e=1669232658&token=IAM-gs8ue1pDIGwtR1CR0Zjdagg7Q2tn5G_1BqTmhmqa:dP1EgsK44tTMjd_a_2XoHiL5v2Y=
倒三叶草形
cover1743_20221122081441.jpg?e=1669128360&token=IAM-gs8ue1pDIGwtR1CR0Zjdagg7Q2tn5G_1BqTmhmqa:uLrYKdsils7UFYZjULKqiN7GS3I=