导图社区 初中英语 词法基础知识 名词
初中英语之词法基础知识——名词思维导图,包括:名词的数、名次所有格、名词的分类。
本章思维导图是关于数据模型的内容,欢迎大家参考。
这是一张关于独处时必看106部高分影片的思维导图,可以看看。
这是一张运营攻略思维导图,内容详尽实用。
社区模板帮助中心,点此进入>>
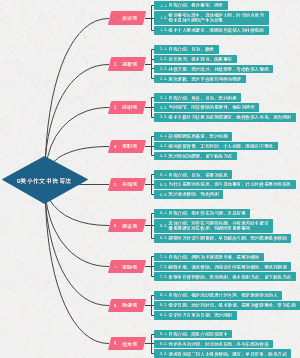
英文雅思大作文的命题类型和写作框架总结!
G类小作文书信写法
六级高频单词
英语学习:P的逻辑
英语学习p字母相关的单词
医学相关的英语词汇
L814音乐
系动词be的语法
48个国际音标标准学习
topic
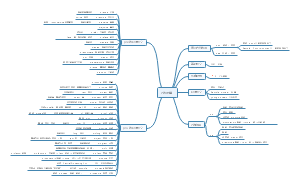
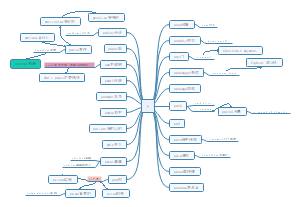
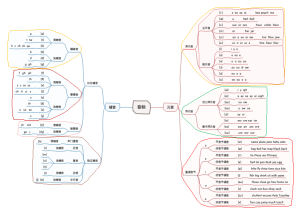
词法基础知识 名词
知识梳理
名词的数
名词单复数的特殊用法
不可数名词的数
可数名词的数
规则变化
不规则变化
名次所有格
's所有格与“名词+of+名词性物主代词”结构
双重所有格
's所有格的用法
's所有个的构成
名词所有格概述
名词的分类
普通名词
不可数名词
抽象名词
物质名词
可数名词
专有名词
学习指导
注意分类
表示人名、地名、书名、节日、组织、机构、星期几、月份等的词。如:Women'sDay、 The United Nations.
不能用数目计算的物质名词或 抽象概念的名词,如: water、 happiness、English等
能用数目计算的名词有单数和 复数之分,如:an apple、two apples 等
注意概念
用以表示人,事物,时间,地点,或抽象概念的名称。
注意”格“
用以表示事务所属关系的刑式
's格
如:Mary's book
of格
如: the name of the book
表示全体中的一部分,为避免重复而既有of格又有's格, 如:one of Tom's friends
总结升华
学习误区
名词作定语
名词作定语时需要用单数形式,但是有 些词例外,如:woman、man作定语时 的形式要随主体词的变化而变化; 作sport作定语时要用复数形式sports
知能提升
常考节日名称
中国节日
3月8日妇女节 Women'sDay
9月10日教师节 Teacher's Day
6月1日儿童节 Children's Day
5月1日劳动节 May Day
10月1日国庆节 the National Day
1月1日元旦 New Year
农历春节 the Spring Festival
外国节日
2月14日情人节 St. Valentine's Day
4月1日愚人节 April Fool's Day
5月的第二个星期日 Mother's Day
6月的第三个星期日 Father's Day
11月的第四个星期日 Thanksgiving Day
12月25日圣诞节 Christmas Day
口诀与归类
口诀
可数名词变复数加s后的发音:清后清, 浊后浊,元音跟着浊音浊。(s如加在清辅 音之后就发清辅音/s/,加在浊辅音之后就 发浊辅音/z/
归类
特列
以o结尾的加es特例:hero-heroes Negro-Negroes