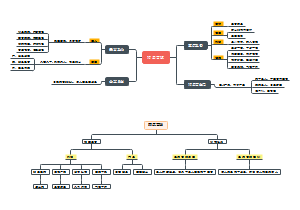
导图社区 高中政治经济生活
高中政治经济生活
这是一个关于高中政治经济生活的思维导图,将知识点进行了归纳和整理,帮助学习者理解和记忆。直击重点,可以作为学习笔记和复习资料,帮助大家系统地回顾和巩固所学知识,知识点系统且全面,希望对大家有所帮助!
编辑于2024-07-12 14:22:07- 经济生活
他的近期作品
查看更多>>
- 高中政治法律与生活
这是一个关于高中政治法律与生活的思维导图,将知识点进行了归纳和整理,帮助学习者理解和记忆。直击重点,可以作为学习笔记和复习资料,帮助大家系统地回顾和巩固所学知识,知识点系统且全面,希望对大家有所帮助!
- 高中政治逻辑与思维-逻辑部分
这是一个关于高中政治逻辑与思维-逻辑部分思维导图,将知识点进行了归纳和整理,帮助学习者理解和记忆。直击重点,可以作为学习笔记和复习资料,帮助大家系统地回顾和巩固所学知识,知识点系统且全面,希望对大家有所帮助!
- 高中政治政治生活
这是一个关于高中政治 政治生活的思维导图,将知识点进行了归纳和整理,帮助学习者理解和记忆。直击重点,可以作为学习笔记和复习资料,帮助大家系统地回顾和巩固所学知识,知识点系统且全面,希望对大家有所帮助!
高中政治经济生活
社区模板帮助中心,点此进入>>
他的近期作品
查看更多>>
- 高中政治法律与生活
这是一个关于高中政治法律与生活的思维导图,将知识点进行了归纳和整理,帮助学习者理解和记忆。直击重点,可以作为学习笔记和复习资料,帮助大家系统地回顾和巩固所学知识,知识点系统且全面,希望对大家有所帮助!
- 高中政治逻辑与思维-逻辑部分
这是一个关于高中政治逻辑与思维-逻辑部分思维导图,将知识点进行了归纳和整理,帮助学习者理解和记忆。直击重点,可以作为学习笔记和复习资料,帮助大家系统地回顾和巩固所学知识,知识点系统且全面,希望对大家有所帮助!
- 高中政治政治生活
这是一个关于高中政治 政治生活的思维导图,将知识点进行了归纳和整理,帮助学习者理解和记忆。直击重点,可以作为学习笔记和复习资料,帮助大家系统地回顾和巩固所学知识,知识点系统且全面,希望对大家有所帮助!
- 相似推荐
- 大纲